
Субъективные и объективные предпосылки возвышения Москвы 🤓 [Есть ответ]
Говоря о субъективных предпосылках возвышения Москвы, следует отдельно отметить политику первых московских князей. Первые два московских князя, Даниил Александрович и Юрий Данилович, присоединили территории по течению Москвы-реки, а также города Коломна и Можайск, Переяславль-Залесский. Стремление Юрия Даниловича стать обособленным князем стала причиной борьбы за Владимир с тверским князем Михаилом Ярославичем. Борьба между тверскими и московскими князьями велась не только за территорию, но из-за ордынский ярлык на великое княжение.
Другим субъективным фактором возвышения Москвы стала политика Ивана Калиты, при которой даже Владимирский митрополит Пётр переехал в Москву. Стараниями Калиты Москва стала центром духовной жизни Восточной Европы, из провинциального города превратилась в центр собирания русских земель.
Объективные факторы – это ряд географических, демографических, экономических и политических предпосылок:
- Москва была провинциальным городом, географически она была удалена от набегов татаро-монголов, при этом здесь скрещивались важные европейские и азиатские торговые пути.

- Из-за спокойствия и размеренного развития московских земель сюда стремились переселиться люди с других городов Древней Руси.
- На территорию Московского княжества, после удачных военных и политических действий московских князей, а также после переноса православный кафедры из Владимира в Москву при Иване Калите, потянулись представители элитных сословий Руси: духовенство, купцы и торговцы, горожане и ремесленники, зажиточные крестьяне, вольные слуги других княжеств и бояре. За счёт последних укреплялась Московская рать.
- В 1367 году началось строительство белокаменного Кремля. Он стал не только неприступной крепостью, но и символом могущества Москвы.
- Не менее важной была поддержка православного духовенства. Вскоре после переноса кафедры митрополит Пётр скончался.
 На его место пришел Феогност, окончательно поселившийся в Москве. На месте постоянного пребывания митрополита начало организовываться монашество.
На его место пришел Феогност, окончательно поселившийся в Москве. На месте постоянного пребывания митрополита начало организовываться монашество. - Таким образом, тогдашнее население Руси могло убедиться в том, что московские князья занимались собирательством земель под предводительством православной церкви.
Причины возвышения Московского княжества
/ /
Причины возвышения Московского княжестваОсновные причины возвышения Московского княжества
Возвышением Московского княжества историки называют процесс роста политической власти и экономического влияния Москвы на остальные княжества, с последующим их объединением Руси вокруг новой столицы.
Впервые Москва упоминается в летописях 1147 года, как город,
возведённый князем Юрием Долгоруким. Изначально Москву называли
Кучково, Москов и представляла она собой крепость, построенную на границе
(южной) Суздальского княжества, которая должна защищать от набегов со стороны
Рязани и Чернигова. Во время монголо-татарского ига город был частично разрушен
и разорён ханом Батыем, продвигающимся от государственных границ к Владимиру и
Суздалю. Как самостоятельная территория Московское княжество начало своё
существование в 1283 году.
Во время монголо-татарского ига город был частично разрушен
и разорён ханом Батыем, продвигающимся от государственных границ к Владимиру и
Суздалю. Как самостоятельная территория Московское княжество начало своё
существование в 1283 году.
Выгодное географическое положение как причина возвышения Москвы
Московское княжество имело очень выгодное географическое положение, располагаясь на перекрёстке путей, которые вели из южной части государственных земель в северную, а также в Рязань из Новгорода. Данное расположение давало возможность городу вести активную торговлю, что давало огромные возможности для политического влияния и экономического роста княжества. Кроме этого, такое выгодное положение привлекало новых жителей, что тоже способствовало приросту экономики области.
Немаловажной причиной экономического и торгового роста было
расположение Москвы на берегу Волги, по водной артерии которой из Рязани в
Новгород перевозили воск и мёд. Таким образом, располагая огромными средствами,
московские князья могли принимать участие в политике государства, диктуя другим
княжествам собственные условия.
Таким образом, располагая огромными средствами,
московские князья могли принимать участие в политике государства, диктуя другим
княжествам собственные условия.
Изобилие денежных средств всё же не главная причина возвышения Московского княжества. Большую роль здесь играло правильное управление первых князей Москвы.
Два первых князя (Даниил Александрович и Юрий Данилович) смогли достаточно увеличить княжеские территории, отобрав у соседей все города по течению Москвы-реки.
После достижения княжеством небывалых высот, Юрий Данилович отправился за ярлыком княжения в Золотую Орду, что впрочем не увенчалось успехом. После его смерти Москвой начал править брат Юрия Иван, который получил прозвище Калита. В отличие от своего брата ему удалось получить от Орды ярлык на княжение. С того времени Московское княжество не упускало никогда своего права.
Рост и возвышение Московского княжества (карта-схема)
Основные причины возвышения Москвы (таблица-схема):
Интересные материалы:
§ 42.
 Причины возвышения Москвы. Учебник русской истории
Причины возвышения Москвы. Учебник русской истории§ 42. Причины возвышения Москвы
Было уже сказано (§ 36), что в эпоху татарского завоевания во Владимиро-Суздальской Руси развился удельный порядок княжеского владения, прекратившийся с усилением Московского княжества.
Особое Московское княжество начало существовать со второй половины XIII в. Самый же город Москва, по преданию, был основан князем Юрием Долгоруким. Летописи упоминают о Москве в первый раз в 1147 г. (затем в 1156 и 1176 гг., называя ее «Москов», «Кучково» и «Москва»). Первоначально Москва была крепостью, поставленной на южной границе Суздальской земли для того, чтобы охранять ее с юга, от Рязани и Чернигова. Батый, идя из Рязани к Суздалю и Владимиру, прежде всего взял и разорил Москву, закрывавшую ему путь к этим городам. В первое столетие своего существования Москва не составляла особого княжества, а принадлежала владимирским князьям. Князь Александр Невский оставил на Москве своего младшего сына Даниила, и только с тех пор Москва стала особым уделом Даниилова потомства. С тех же пор началось быстрое усиление и возвышение нового удела.
С тех же пор началось быстрое усиление и возвышение нового удела.
А. Васнецов. Постройка новых стен Кремля Юрием Долгоруким в 1156 году
Первая причина возвышения Москвы и усиления Московских князей заключалась в особенно выгодном географическом положении города Москвы. Москва была расположена в узле дорог, ведших из южной Руси в северную и из Новгородской земли в Рязанскую. Переселенцы из южных областей, идя на север, не могли миновать реки Москвы и в большом числе оседали вокруг города Москвы ранее, чем подвигались далее к северу. Оттого Московское княжество быстро и густо заселялось, а это давало московским князьям большие средства, так как чем гуще было население удела, тем больше получал удельный князь. Река Москва, с другой стороны, была таким водным путем, который соединял верховья Волги со средним течением Оки. Этим путем пользовались новгородцы, перевозя к себе хлеб, воск и мед из богатейших мест Рязанской земли.
А. Васнецов. Строительство деревянных стен Кремля. XII век
Второю причиною возвышения Москвы были таланты и ловкость первых московских князей, умевших использовать выгоды своего положения. Первые два московских князя, Даниил Александрович и сын его Юрий, успели «примыслить» себе все течение Москвы-реки, отняв от Рязанского князя город Коломну на устье р. Москвы и от Смоленского князя город Можайск на верховьях р. Москвы. Кроме того, князь Даниил получил город Переяславль-Залесский по завещанию бездетного Переяславского князя.
Данный текст является ознакомительным фрагментом.
Предпосылки объединения русских земель возвышение москвы
ПЛАН-КОНСПЕКТ УРОКА
Предпосылки объединения русских земель. Возвышение Москвы.
Ф.И.О.: Осипова Любовь Ивановна
Место работы: Муниципальное общеобразовательное
учреждение
средняя общеобразовательная школа №3
с углубленным изучением отдельных предметов
Должность: учитель
Предмет: история
Класс: 10
6. Тема
и номер урока в теме: Объединение
русских земель. №1.
Тема
и номер урока в теме: Объединение
русских земель. №1.
7. Базовый учебник: Учебник: А.Н. Сахаров, В.И.Буганов.
«История России с древнейших времен
до конца ХVII в.». М., «Просвещение»,2003.
8. Цель урока: раскрыть предпосылки объединения русских земель
в единое государство, показать закономерность
процесса образования централизованного государства.
9. Задачи:
Образовательные: формировать представление о
процессе централизации русских земель,
о причинах возвышения Москвы. Формировать
представления у учащихся о роли личности
в истории на примере деятельности
московских князей.
Формировать
представления у учащихся о роли личности
в истории на примере деятельности
московских князей.
развивающие: — продолжить работу над развитием у учащихся навыков деятельности с исторической картой, историческими источниками и ЭОР на уроках и во внеурочное время;
-формировать умение анализировать, сопоставлять исторические события, делать самостоятельно исторические выводы.
воспитательные: воспитывать чувство патриотизма и любви к Родине
на примере героизма народов России в борьбе с завоевателями.
10. Тип урока: урок изучения нового материала.
11. Формы работы учащихся: фронтальная, индивидуальная, коллективная.
12. Необходимое техническое оборудование: компьютер, мультимедийный проектор, ЭОР с сайта ФЦИОР, ЕКЦОР.
Таблица 1
СТРУКТУРА И ХОД УРОКА
№ п/п | Этап урока | Название используемых ЭОР (с указанием порядкового номера из таблицы 2) | Деятельность учителя (с указанием действий с ЭОР, например, демонстрации) | Деятельность ученика | Время в мин |
1 | 2 | 3 | 4 | 5 | 6 |
1 | Вступитель- ное слово учителя | Объявляет
тему и цель урока. | Запись даты, темы урока | 2 | |
2 | Проверка домашнего задания | Фронтальный опрос по итогам выполнения домашнего задания по теме: «Главные политические центры Руси в период феодальной раздробленности» (отвечает на возникающие у учащихся вопросы. | Отвечают на вопросы,используя самостоятельно составленные схемы «Особенности цивилизацион- ных моделей Киевской Руси» | 5 | |
3 | Мотивационная заставка. Актуализация полученных знаний учащихся | Постановка проблемы в форме ряда вопросов: 1132г. Новгородский летописец записал »И
раздрася вся земля. -О каком периоде истории русского государства повествует летописец? -Что послужило предпосылкой объединения русских земель? -Какой фактор в объединении русских земель был определяющим в сравнении с Европой? -Каковы возможные центры объединения? Проблемный вопрос на урок. -Почему именно Москва становится центром объединения? | Ответы на проблемные вопросы совместно с учителем. Формулирование темы урока. | 7 | |
4 | Изучение нового материала | №1. «Предпосылки объединения русских земель» №2. «Карта Московского княжества XIVв.» №3.«Схема «Возвышение Москвы» №8.»Потомки Александра Невского» №4. №5.»Подавление антиордынского восстания в Твери.Миниатюра из летописного лицевого свода» №7. «Духовная грамота Ивана Калиты» | Объясняет новый материал, используя материалы ЭОР как основу для презентации: 1.Предпосылки объединения русских земель. -Укрепление русской государственности. -Возрождение экономики русских княжеств. — Потеря Киевом своей значимости. -Необходимость освобождения от монголо- татар. 2. Возможные центры объединения. Северо- Западная Русь. Русь Литовская. Северо- Восточная Русь. 3.Причины возвышения Москвы 4.Правление Ивана Калиты и его наследников. | Воспринимают
информацию, сообщаемую учителем,
составляют план в тетради «Предпосылки
объединения русских земель» работают
с картой. Учащиеся отвечают на вопрос: почему ни Северо- Западная, ни Русь Литовская не имела шансов на то, чтобы стать центром объединения? Работа со схемой Составление плана «Причины возвышения Москвы» (примерный план) 1. Удобное географическое положение. 2. Сочувствие духовенства и боярства. 3.Политическая близорукость татар. 4.Личные способности московских князей. Работа в группах с документами и иллюстрациями. Дают оценку деятельности Ивана Калиты и его наследников. | |
5 | Закрепление и обобщение изученного материала | №9. «Контрольные вопросы» | Учитель инструктирует учащихся по выполнению заданий на основе ЭОР. | Выполняют
тест в тетради с последующей проверкой
на слайде. | 6 |
6 | Домашнее задание | Инструктаж по выполнению домашнего задания (параграф 22,В.и.22(п.1-2) вопросы и задания к нему).Подготовить сообщения:»Сергий Радонежский- воплощение национального нравственного идеала», «Роль русской православной церкви в объединении Руси» (по желанию учащихся) | Слушают инструктаж, записывают задание в дневники. | 2 | |
7 | Итог урока | №9.»Аудифрагмент. Воспоминание иностранцев о Москве» | Подводит итоги урока, ставит обобщающие вопросы. Почему именно Москва становится центром объединения русских земель? Было ли это случайностью, капризом истории или исторической неизбежностью? | Отвечают
на поставленные вопросы и делают вывод
по уроку. | 5 |

 »
» »Иван
Калита»
»Иван
Калита»

 Обсуждают и оценивают свою
деятельность на уроке.
Обсуждают и оценивают свою
деятельность на уроке.Приложение к плану-конспекту урока
Начало объединения русских земель.
Возвышение Москвы.
Таблица 2
ПЕРЕЧЕНЬ ИСПОЛЬЗУЕМЫХ НА ДАННОМ УРОКЕ ЭОР
№ | Название ресурса | Тип, вид ресурса | Форма предъявления информации (иллюстрация, презентация, видеофрагмен-ты, тест, модель и т.д.) | Гиперссылка на ресурс, обеспечивающий доступ к ЭОР |
1 | «Предпосылки объединения русских земель» | Информационный
тип. Демонстрационный материал. | Справочный материал с иллюстрациями | Предпосылки объединения русских земель Духовная грамота Ивана Калиты Контрольные вопросы. Потомки Александра Невского |
2 | «Карта .Московское княжество в начале XIVв.» | Информационный тип. Демонстрационный материал. | Карта | Карта «Московское княжество в начале XIV в.» |
3 | «Схема. Возвышение Москвы» | Информационный тип. Демонстрационный материал. | Анимированная интегративная схема. | Схема: «Возвышение Москвы» |
4 | «Иван Данилович Калита» | Информационный
тип. Демонстрационный материал. | Иллюстрация | Иван Данилович Калита |
5. | «Подавление антиордынского восстания в Твери. Миниатюра из лицевого летописного свода» | Практический тип | Документ | Подавление антиордынского восстания в Твери. Миниатюра из лицевого летописного свода |
6 | «Аудифрагмент Воспоминания иностранцев о Москве». | Информационный тип | Аудифрагмент | Аудиофрагмент: воспоминания иностранцев о Москве |
7 | «Духовная грамота Ивана Калиты» | Практический тип | Документ | Духовная грамота Ивана Калиты |
8 | «Потомки Александра Невского» | Информационный тип. Демонстрационный материал | Анимированная интегративная схема. | Потомки Александра Невского |
9 | «Контрольные вопросы» | Контрольный тип. Тренировочно- контролирующий материал | Интерактивное задание | Контрольные вопросы. |



Возвышение Москвы
Установление единодержавия в Московской Руси и возвышение значения великокняжеской власти
28.10.2008/реферат
Общий характер собирательной деятельности московских князей. Иван III, установление единодержавия. Отстройка резиденции великого князя. Новые титулы великого князя и венчание на великое княжение. Содействие духовенства возвеличению Московского государя.
Причины возвышения Москвы30.11.2011/контрольная работа
Возвышение Москвы. . Предпосылки возвышения Москвы Политическая линия Московский князей. Схватка Твери и Москвы Окончательная централизация власти вокруг Москвы Историческая личность. Иван Калита.
. Предпосылки возвышения Москвы Политическая линия Московский князей. Схватка Твери и Москвы Окончательная централизация власти вокруг Москвы Историческая личность. Иван Калита.
4.05.2018/реферат
Формирование и возвышение Московского государства (XIII—XV вв.)
А.Д. Меншиков как государственный деятель6.12.2008/курсовая работа
Меншиков — государственный деятель Российской империи. Вопрос о происхождении Меншикова. Деятельность Меншикова в период Северной войны. Возвышение не политической арене. Роль Меншикова во внутренних преобразованиях Петра Великого. Потеря власти и ссылка.
Борис Годунов — реформатор России10.02.2010/реферат
Историческая обстановка в России конца XVI века. Сыновья Ивана IV (Грозного) — претенденты на власть, род Годуновых. Характер Бориса Годунова и его возвышение, реформаторский характер правления при Федоре Ивановиче. Избрание Бориса на царство и смута.
Борьба за власть в советском руководстве 1946-1953 г.
18.12.2007/реферат
Перемены в структуре аппарата ЦК и обновление их руководства в период с 1945 г. до смерти Сталина. Работа обновлённого ЦК и одновременная борьба за власть. Смерть Сталина и возвышение Хрущева открыла новый период в истории страны-«хрущевская оттепель».
Возвышение Москвы и образование Российского государства12.05.2009/дипломная работа
Москва как самый большой мегаполис в мире. Причины и характеристика основных этапов возвышения Москвы, взгляды ведущих историков на этот процесс. Московское княжество в правлении Даниила Александровича Московского в период наибольшего дробления Руси.
Возвышение Москвы и объединение вокруг нее северо-восточной Руси28.10.2008/реферат
Подготовка нового государственного порядка. Собирательная деятельность московских князей. Причины возвышения Москвы. Прилив населения в бассейн реки Москвы. Содействие ханов Золотой орды и общественных сил. Противодействие со стороны других княжеств.
Древний Египет18. 01.2010/реферат
01.2010/реферат
Египет в эпоху раннего и древнего царства, образование развитых неолитических общин. Объединение долины Нила в единое государство. Период Среднего царства, деление Египта, набеги ливийцев и кочевников. Эпоха нового царства и возвышения фиванских царей.
Начало объединения русских земель вокруг Москвы7.10.2004/реферат
Первое упоминание о Москве и причины её возвышения. Географическое положение Москвы. Политическая деятельность первых московских князей. Влияние православной церкви на возвышение Москвы. Междуусобица в первой половине XV века.
Специфика формирования единого российского государства
| Объективные причины | Субъективные причины | Случайные причины |
|---|---|---|
| 1) Географическое положение 2) Переселение людей в Москву 3) Поддержка идеи объединения Руси массами.  | 1) Политика князей 2) Деятельность церкви | 1) Поддержка Ордой 2) Чума в Москве |
Блок: 1/5 | Кол-во символов: 370
Источник: https://istoriarusi.ru/udel/pricini-vozvisheniya-moskvi-14-vek.html
Возрождение из руин как предпосылка единения Руси
Хозяйственный, ремесленный и торговый упадок, охвативший Русь после вторжения Батыя, казалось бы, навсегда похоронил надежду на возрождение единого государства. Главной специфической особенностью складывания предпосылок для создания централизованной русской державы были тяжелейшие условия ордынского ига.
Блок: 2/5 | Кол-во символов: 354
Источник: https://www.istmira.com/drugoe-istoriya-rossii/13390-specifika-formirovaniya-edinogo-rossiyskogo-gosudarstva.html
Объективные причины
Главные объективные причины возвышения Москвы над другими городами и княжествами Удельной Руси:
- Удобное географическое положение. Город был относительно удален от набегов монголов, а также здесь перекрещивались важные торговые пути.

- Увеличение численности населения. Люди старались переселиться в эти земли, как более удобные для жизни.
- Большинство населения поддерживало идеи объединения Руси.
Далее мы рассмотрим основные особенности важнейших причин возвышения Московского княжества, но вначале необходимо отметить, что важную роль в этом процессе сыграла поддержка населения. За объединение Руси в единое государство, вокруг единого политического, экономического и духовного центра выступали: бояре (в основном служилые), дворяне, духовенство, купечество, ремесленники и крестьяне. То есть — основная масса населения была «за» осуществление этих процессов.
Если кратко рассматривать причины возвышения Москвы в 14 веке, то все существующие аргументы можно разделить на 2 большие группы: объективные и субъективные факторы.
Географические причины
Если посмотреть на карту Руси и Московского княжества в 13-14 веках, то становится понятным почему именно географические причины возвышения Москвы принято считать главенствующими. Существуют 2 географические причины возвышения Московского княжества:
Существуют 2 географические причины возвышения Московского княжества:
- Удаленность от Золотой Орды.
- Пересечение важных торговых путей
Через Москву проходил торговый путь из Новгорода в Смоленск (2 самых богатых торговых городов Руси), а также торговый путь из Оки на Волгу, откуда торговцы добирались в Золотую Орду. Пересечение 2-ух крупных торговых путей позволяло Москве жить относительно безбедно только за счет установления пошлины.
Красным на карте обозначены изначальные границы Москвовского княжества.
Блок: 2/5 | Кол-во символов: 1803
Источник: https://istoriarusi.ru/udel/pricini-vozvisheniya-moskvi-14-vek.html
Лидеры и стяжатели централизации Руси
Одержав верх в борьбе за первенство в объединительном процессе, московские князья сосредоточили в своих руках политическую власть в северо-восточных землях. Приняв у себя митрополита, Москва обрела положение духовного центра всей Руси. Здесь постепенно складывалось территориальное ядро единого государства, и формировалась его политическая основа.
В централизации русских земель выгоду приобретали дворяне, купеческое сословие и ремесленники, возлагавшие надежды на упрочение экономических связей. Феодалы хотели использовать силу единого государства в дальнейшем юридическом закабалении крестьянства. Все слои населения Руси нуждались в сильном государстве, способном отразить агрессию внешнего врага.
Блок: 3/5 | Кол-во символов: 741
Источник: https://www.istmira.com/drugoe-istoriya-rossii/13390-specifika-formirovaniya-edinogo-rossiyskogo-gosudarstva.html
Субъективные причины
Выше мы рассмотрели объективные причины, а субъективные причины возвышения Москвы над другими княжествами в 14 веке следующие:
- Политика первых князей. Московские князья сразу начали вести политику на собирание земель.
- Церковь. После того как митрополит Петр перебрался из Владимира в Москву, именно Москва стала религиозным центром Руси.
Политика князей
Политика местных князей строилась на получении независимости, а также постепенного подчинения себе соседей. И рост территории действительно начался. Достаточно посмотреть на карту.
И рост территории действительно начался. Достаточно посмотреть на карту.
Карта — Рост территории Московского княжества в 14 веке
Роль церкви
При Иване Калите митрополит Владимирский Петр переехал в Москву. Город моментально стал центром духовной жизни Руси. при этом церковь на тот момент уже высказывала идеи необходимости объединения Руси вокруг единого политического центра. Естественно, что в глазах церкви и в ее учениях московские князья стали правителями всех русских земель. Этим воспользовался Иван Калита, который объявил себя князем Московским и всея Руси.
Блок: 3/5 | Кол-во символов: 1078
Источник: https://istoriarusi.ru/udel/pricini-vozvisheniya-moskvi-14-vek.html
Факторы внешней политики
Монголо-татарское нашествие стабилизировало феодальное раздробление Руси и одновременно ослабило всесторонние связи между разными регионами страны. Путь «из варяг в греки» совсем потерял актуальность. Новый внешнеполитический вектор «запад — восток», переместился на русский северо-восток. Политика Александра Невского и православной церкви пошла по пути дистанцирования от духовной агрессии Запада в сторону веротерпимой Орды. Этот вектор положил начало складыванию великорусской народности.
Политика Александра Невского и православной церкви пошла по пути дистанцирования от духовной агрессии Запада в сторону веротерпимой Орды. Этот вектор положил начало складыванию великорусской народности.
В XIII — XV веках, для политической централизации сложились более благоприятные условия, чем для экономической. Этому немало способствовало наличие внешней угрозы как с Запада, так и с Востока. Борьба за освобождение от золотоордынского ига и установление национального суверенитета катализировали объединительные процессы.
Блок: 4/5 | Кол-во символов: 841
Источник: https://www.istmira.com/drugoe-istoriya-rossii/13390-specifika-formirovaniya-edinogo-rossiyskogo-gosudarstva.html
Случайные факторы
В большинстве учебников приводится огромное количество причин и факторов объединения земель вокруг Москвы в 14 веке, но при этом абсолютно игнорируется факт того, что в этом процессе были случайные факторы, которые нельзя отнести ни к объективным, ни к субъективным. При этом эти факторы были крайне важны. Например, один из случайных факторов — эпидемия чумы в Московском княжестве при правлении Ивана Калиты. Из-за чумы погибло большинство детей и внуков князя. В живых остались только 2 внука: Дмитрий Иванович и Владимир Андреевич. Этот факт позволил избежать Москве суровых междоусобных войн, а также дробления княжества на мелкие удельные куски.
При этом эти факторы были крайне важны. Например, один из случайных факторов — эпидемия чумы в Московском княжестве при правлении Ивана Калиты. Из-за чумы погибло большинство детей и внуков князя. В живых остались только 2 внука: Дмитрий Иванович и Владимир Андреевич. Этот факт позволил избежать Москве суровых междоусобных войн, а также дробления княжества на мелкие удельные куски.
Во-многом к случайным факторам можно отнести и покровительство ордынских ханов над Москвой. Они неосмотрительно делали Москву центром, не полагая, что небольшой город может представлять собой большую угрозу. Например, ханы наделили московских князей правом собирать для них дань.
Блок: 4/5 | Кол-во символов: 969
Источник: https://istoriarusi.ru/udel/pricini-vozvisheniya-moskvi-14-vek.html
Специфические отличия процесса централизации Руси
Формирование русского единого государства отличалось от аналогичных процессов в Европе тем, что:
1. Объединение Руси вершилось в эпоху позднего феодализма, тогда как Европа централизовалась в период его расцвета.
2. Собиранием доменов под свою руку занимались московские князья, а в европейских странах, мелкопоместное дворянство и городская буржуазия.
3. Политические причины доминировали в процессе централизации Руси, а в Европе экономические.
Завершилось формирование единого государства в XVI веке, с воцарением Ивана Грозного, поставившего под контроль своих наместников всех удельных князей.
Таким образом, главной специфической чертой создания русского государства было то, что политическая централизация открыла путь для экономического единения земель крупнейшего государства в Европе. Образование полноценного государства стало возможным только в княжествах, зависимых от Золотой Орды.
Блок: 5/5 | Кол-во символов: 953
Источник: https://www.istmira.com/drugoe-istoriya-rossii/13390-specifika-formirovaniya-edinogo-rossiyskogo-gosudarstva.html
Количество использованных доноров: 3
Информация по каждому донору:
- https://www.
 istmira.com/drugoe-istoriya-rossii/13390-specifika-formirovaniya-edinogo-rossiyskogo-gosudarstva.html: использовано 4 блоков из 5, кол-во символов 2889 (27%)
istmira.com/drugoe-istoriya-rossii/13390-specifika-formirovaniya-edinogo-rossiyskogo-gosudarstva.html: использовано 4 блоков из 5, кол-во символов 2889 (27%) - https://histerl.ru/lectures/moskovskaia_rusi/predposilki_obiedinenia_russkix_zemeli.htm: использовано 1 блоков из 3, кол-во символов 3421 (32%)
- https://istoriarusi.ru/udel/pricini-vozvisheniya-moskvi-14-vek.html: использовано 4 блоков из 5, кол-во символов 4220 (40%)
Собирание русских земель во второй половине XIV в. Куликовская битва
Слайд 1
Цель урока: раскрыть значение Куликовской битвы в истории России.
Основные знания: дата Куликовской битвы, участники, ее этапы и историческое значение.
Работа с исторической картой: показать на карте место Куликовской битвы.
Работа с документами: сравнительная таблица о готовности к битве Руси и Золотой Орды, фрагмент из «Жития Сергия Радонежского»
Основные персоналии: Дмитрий Донской, Сергий Радонежский, Мамай.
Основная дата: 8 сентября 1380 г. – Куликовская битва.
Опережающее задание: подготовить сообщения о Дмитрии Донском и Преподобном Сергие Радонежском.
Межпредметные и внутрикурсовые связи: история России: причины возвышения Москвы (§18 учебника).
Образовательные результаты: развитие личностных и метапредметных УУД.
Оборудование урока: ПК(+колонки), проектор, экран.
План изучения нового материала:
- Москва – центр объединения северо-восточных русских земель.
- Русь готовится к борьбе за свободу.
- Поход Мамая на Русь.
- Битва на поле Куликовом.
- Значение Куликовской битвы.
Слайд 2
Перед вами три этапа в результате которых Москва становилась центром Древнерусского государства. На предыдущих уроках мы познакомились с первым этапом возвышения Москвы. Мы узнали с вами как, начиная со второй половины 13 века и до начала 14-го происходило формирование таких крупных центров как Тверь, Москва и Рязань. И каким образом из этих городов Москва начала возвышаться.
Мы узнали с вами как, начиная со второй половины 13 века и до начала 14-го происходило формирование таких крупных центров как Тверь, Москва и Рязань. И каким образом из этих городов Москва начала возвышаться.
Давайте вспомним с вами причины возвышения Москвы и предпосылки объединения русских земель.
Слайд 3
1. Расставьте причины возвышения Москвы в порядке их значимости. (Порядковые номера записать в сравнительной таблице на доске):
- А) Удобство географического положения.
- Б) Расположение на перекрестке торговых путей.
- В) Умелая политика московских князей.
- Г) Поддержка московских князей православной церковью.
- Д) Москве удалось избежать дробления из-за того, что в княжеских землях было мало сыновей.
- Е) Поддержка московских князей ордынскими ханами.
Слайд 4
2. Отметьте в первой колонке таблицы (на доске) причины, а во второй – предпосылки (предварительные условия) объединения русских земель.
- Хозяйственное возрождение Северо-Восточной Руси.
- Восстановление торговых связей между княжествами.
- Необходимость свержения ордынского владычества.
- Сохранение единства церковной организации.
- Единая вера, язык и культурные традиции.
Слайд 5
Перед вами появилась тема нашего сегодняшнего урока «Собирание русских земель во второй половине XIV века. Куликовская битва». Зная ее, давайте обозначим цель сегодняшнего урока. О чем мы с вами будем разговаривать? А в конце урока мы определим, достигли ее или нет. И тогда станет ясно, правильная ли мы поставили перед собой цель.
(Предполагаемая цель урока: раскрыть значение Куликовской битвы в объединении русских земель.)
План урока: (Слайд 6)
3. Москва – центр объединения северо-восточных русских земель.Слайд 7
Сегодня мы с вами познакомимся со вторым этапом объединения земель вокруг Москвы и как она встает во главе борьбы с татаро-монгольским игом.
Слайд 8
В 1359 году на московском престоле оказывается девятилетний внук Ивана Калиты Дмитрий Иванович. Конечно, такой маленький ребенок не мог самостоятельно править княжеством. От его имени Москвой управлял митрополит Алексей (Клик), сын знатного московского боярина. Он приложил немало усилий к тому, чтобы Орда признала Дмитрия, не по годам развитого и смышленого мальчика, великим владимирским князем.
Алексей продолжил политику Ивана Калиты по собиранию русских земель вокруг Москвы.
Слайд 9
Московскому княжеству удалось подчинить себе главных соперников – суздальско-нижегородского, тверского и рязанского князей, а также отбить три похода литовцев на Москву.
Слайд 10
И вот Дмитрий, достигнув совершеннолетия начинает вести такую же энергичную и смелую политику как и его дед. Продолжает расширять пределы своего княжества, включив в состав своих владений Углич, Галич и Белоозеро и приобретя ряд новых городов с их округами… Он ведет успешную борьбу со своими соперниками, великими князьями тверскими и рязанскими, которые заключили опасный союз с противником Москвы великим князем Литовским Ольгердом…
Но главной целью политики Дмитрия было объединение Руси.
А теперь давайте послушаем сообщение Вити о Дмитрии Донском. После его сообщения запишем в тетради годы правления Дмитрия Донского 1359-1389 гг. Слушаем внимательно.
(По ходу рассказа идет презентация о Донском)
4. Русь готовится к борьбе за свободу.Слайд 11
| Золотая Орда | Русские земли |
|
|

Слайд 12
У вас на столах документы. Внимательно изучите их и скажите мне: Какая из сторон была более готова к началу Куликовской битвы. Ответ обоснуйте.
(Ответ учащихся: Русские земли были сильнее, потому что происходило их объединение, а Орда становил раздробленным государством).
5. Поход Мамая на Русь.
Поход Мамая на Русь.Вы уже узнали о том, что в 1378 году состоялась битва с ордынцами на реке Вожа. Это сражение впервые поставило под сомнение власть Орды над Русью.
Слайд 13
-Ребята сейчас мы рассмотрим с вами положение Орды и Руси накануне Куликовской битвы:
Мамай не смог смириться с поражением. Это означало бы признание независимости Руси и потерю большой дани, которую выплачивали княжества, поэтому обе стороны начинают подготовку к решающему сражению.
Слайд 14
Как видно на экране готовность армии с каждой стороны была практически одинаковой.
Но, прежде, чем принято бой с татарами, Дмитрий обратился за благословением к преподобному Сергию Радонежскому. И тот вместе с благословением отправил с князем монахов-богатырей Пересвета и Ослабю, чтобы показать, чо дело освобождения родины свято даже для иноков, покинувших мирскую жизнь.
Что это за человек Сергий Радонежский и почему Дмитрий пришел к нему за благословением мы узнаем из сообщения Маши. (Маша рассказывает о Сергии Радонежском, опираясь на слайды презентации).
(Маша рассказывает о Сергии Радонежском, опираясь на слайды презентации).
И вот 6 сентября 1380 года войско Дмитрий Донского двинулось в путь… Внимательно посмотрите фильм о битве на поле Куликовом и после просмотра мы с вами запишем дату Куликовской битвы и ее итог в тетрадь. (Показывается фильм о Куликовской битве).
Слайд 15
Фильм закончился и мы запишем с вами в тетрадь дату битвы.
Когда же состоялась Куликовская битва? (8 сентября 1380 года)
Слайд 16
Итог битвы? (Мамай со своим войском сбежал с поля боя).
Слайд 17
Посмотрев фильм, мы познакомились с историческим итогом битвы. Но существует еще и иная версия. С ней вы познакомитесь, прочитав фрагмент из документа «Житие Сергия Радонежского». Внимательно изучите его и скажите: в чем видит автор «Жития» причины победы в поле Куликовом и согласны ли вы с ними? Свой ответ обоснуйте.
Слайд 18
А теперь давайте с вами определим, и запишем в тетрадь: в чем же историческое значение Куликовской битвы? Что дала русским победа в ней?
Слайд 19
1. Куликовская битва стала коренным переломом в борьбе Руси против татаро-монгольского ига;
2. Оказала решающее влияние на создание единого русского государства;
7. Домашнее задание:Слайд 20
- Учебник §19
- Рабочая тетрадь §18, задания 1,3,5.
- Подготовка к семинару «Культура и быт Руси в XIV – XV вв» (деление класса на 4 группы и раздача дополнительного материала с заданиями для групп
Слайд 21
На этом значении Куликовской битвы изучение сегодняшней темы позвольте считать законченной, и напоследок подведем итоги: На доске в начале урока мы поставили для себя цель. Достигли ли мы ее? (да) Значит правильную цель обозначили? (Да). Ребята по кругу высказываются одним предложением, выбирая начало фразы из рефлексивного экрана на доске.
Ребята по кругу высказываются одним предложением, выбирая начало фразы из рефлексивного экрана на доске.
% PDF-1.4
%
447 0 объект
>
эндобдж
xref
447 38
0000000016 00000 н.
0000001129 00000 н.
0000001268 00000 н.
0000001424 00000 н.
0000001576 00000 н.
0000001717 00000 н.
0000002412 00000 н.
0000002813 00000 н.
0000002852 00000 н.
0000003012 00000 н.
0000003171 00000 п.
0000003765 00000 н.
0000003946 00000 н.
0000004153 00000 п.
0000004366 00000 н.
0000004949 00000 н.
0000004979 00000 п.
0000005002 00000 н.
0000006173 00000 п.
0000006196 00000 н.
0000007306 00000 н.
0000007328 00000 н.
0000008390 00000 н.
0000008413 00000 н.
0000009505 00000 н.
0000009528 00000 н.
0000010651 00000 п.
0000010673 00000 п.
0000011752 00000 п.
0000011775 00000 п.
0000012910 00000 п.
0000012933 00000 п.
0000014027 00000 п.
0000014103 00000 п.
0000014170 00000 п.
0000014249 00000 п. \) u \ rs! e ֨ öJ [\ (ЃIY)
/ U (uэb_hseI6Y0 \ rxr)
/ P -60
/ V 1
/ Длина 40
>>
эндобдж
450 0 объект
> / Кодировка> >>
/ DA (? DžT0m1 \\ U)
>>
эндобдж
451 0 объект
>
эндобдж
452 0 объект
>
эндобдж
483 0 объект
>
ручей
-g I {% gC6W8DT (S \ pҥ- (LH «> Sk5 (~ _yy] # q $ Ȓhw [.3 ٻ> cBj2jG (‘США’ BX`t
qOm
LqV0TJF @ ψ7
XF.a? GRgq0X {iIĖ`} 6 ‘_ (yCZljWd
~ l˒tp) GR
конечный поток
эндобдж
484 0 объект
526
эндобдж
453 0 объект
> / ExtGState>
/ Font> / ProcSet [/ PDF / Text] >>
/ Содержание [464 0 R 466 0 R 468 0 R 470 0 R 472 0 R 474 0 R 476 0 R 478 0 R]
/ MediaBox [0 0 612 792]
/ CropBox [0 0 612 792]
/ Повернуть 0
/ StructParents 0
/ Аннотации 454 0 руб.
>>
эндобдж
454 0 объект
[
455 0 руб. 456 0 руб.
]
эндобдж
455 0 объект
>
эндобдж
456 0 объект
>
эндобдж
457 0 объект
>
эндобдж
458 0 объект
[
/ CalRGB> ]
эндобдж
459 0 объект
>
эндобдж
460 0 объект
>
эндобдж
461 0 объект
>
эндобдж
462 0 объект
/ DeviceGray
эндобдж
463 0 объект
1091
эндобдж
464 0 объект
>
ручей
kr} sljQqd # {y} (U] ϶QIE ‘>) Ê`}! k [? ȍJЗ6,8 «sjīGL52
u5V_k-trVĿJSY ߊ \ GpzI0
q} IlEI [R} qz: bH # ʟ / # ^ Jk ׀ M`y &) Fq \ «mTw (4 [5R ‘; 45» gQ @ ҮuE @> кг, ua˿D * SAZm {4yC, EYӨ5CC] знак равно
WoSv ~.
\) u \ rs! e ֨ öJ [\ (ЃIY)
/ U (uэb_hseI6Y0 \ rxr)
/ P -60
/ V 1
/ Длина 40
>>
эндобдж
450 0 объект
> / Кодировка> >>
/ DA (? DžT0m1 \\ U)
>>
эндобдж
451 0 объект
>
эндобдж
452 0 объект
>
эндобдж
483 0 объект
>
ручей
-g I {% gC6W8DT (S \ pҥ- (LH «> Sk5 (~ _yy] # q $ Ȓhw [.3 ٻ> cBj2jG (‘США’ BX`t
qOm
LqV0TJF @ ψ7
XF.a? GRgq0X {iIĖ`} 6 ‘_ (yCZljWd
~ l˒tp) GR
конечный поток
эндобдж
484 0 объект
526
эндобдж
453 0 объект
> / ExtGState>
/ Font> / ProcSet [/ PDF / Text] >>
/ Содержание [464 0 R 466 0 R 468 0 R 470 0 R 472 0 R 474 0 R 476 0 R 478 0 R]
/ MediaBox [0 0 612 792]
/ CropBox [0 0 612 792]
/ Повернуть 0
/ StructParents 0
/ Аннотации 454 0 руб.
>>
эндобдж
454 0 объект
[
455 0 руб. 456 0 руб.
]
эндобдж
455 0 объект
>
эндобдж
456 0 объект
>
эндобдж
457 0 объект
>
эндобдж
458 0 объект
[
/ CalRGB> ]
эндобдж
459 0 объект
>
эндобдж
460 0 объект
>
эндобдж
461 0 объект
>
эндобдж
462 0 объект
/ DeviceGray
эндобдж
463 0 объект
1091
эндобдж
464 0 объект
>
ручей
kr} sljQqd # {y} (U] ϶QIE ‘>) Ê`}! k [? ȍJЗ6,8 «sjīGL52
u5V_k-trVĿJSY ߊ \ GpzI0
q} IlEI [R} qz: bH # ʟ / # ^ Jk ׀ M`y &) Fq \ «mTw (4 [5R ‘; 45» gQ @ ҮuE @> кг, ua˿D * SAZm {4yC, EYӨ5CC] знак равно
WoSv ~. dHsRw6tMu Lrdq8 öV> -Y \ gkjx3VC̄
\ BDžV鿇’C2xBAfkUxZo || uRZc.
dHsRw6tMu Lrdq8 öV> -Y \ gkjx3VC̄
\ BDžV鿇’C2xBAfkUxZo || uRZc.
Изменение высоты поверхности ледников Тоттен и МГУ от …
Контекст 1
… Характер изменения высоты по данным ICESat-1. Мы нашли изменения высоты поверхности по данным GLAS / ICESat-1 за период 2003-2008 гг. И скорректировали их с учетом десятилетней изменчивости глубины фирна (рис. 1a, b). У результатов есть две важные особенности. Во-первых, это высокая скорость опускания ледяной поверхности в заземленных частях ТГ.Другой — заметный контраст между этим понижением поверхности и меньшим изменением, наблюдаемым в области MUG. Самые высокие темпы опускания поверхности на заземленных участках, окружающих ТГ, по-видимому, совпадают с …
Контекст 2
… результаты имеют две характерные особенности. Во-первых, это высокая скорость опускания ледяной поверхности в заземленных частях ТГ. Другой — заметный контраст между этим понижением поверхности и меньшим изменением, наблюдаемым в области MUG. Самые высокие темпы опускания поверхности в заземленных районах, окружающих ТГ, по-видимому, совпадают с наиболее быстрым течением льда (рис.1а, врезка). Таким образом, за 6-летний период наблюдений поверхность опускалась на 80-170 см / год (неопределенность оценивается в Методиках) выше по течению от линии заземления основного быстротечного ствола ледника (точка А. , Рис. 1а). Далее вверх по течению, следуя кривизне ледника вглубь суши, поверхность опускалась медленнее …
Самые высокие темпы опускания поверхности в заземленных районах, окружающих ТГ, по-видимому, совпадают с наиболее быстрым течением льда (рис.1а, врезка). Таким образом, за 6-летний период наблюдений поверхность опускалась на 80-170 см / год (неопределенность оценивается в Методиках) выше по течению от линии заземления основного быстротечного ствола ледника (точка А. , Рис. 1а). Далее вверх по течению, следуя кривизне ледника вглубь суши, поверхность опускалась медленнее …
Контекст 3
… опускания поверхности в мелиорированных областях, окружающих ТГ, похоже, совпадают с наиболее быстрым движением льда ( Инжир.1а, врезка). Таким образом, за 6-летний период наблюдений поверхность опускалась на 80-170 см / год (неопределенность оценивается в Методиках) выше по течению от линии заземления основного быстротечного ствола ледника (точка А. , Рис. 1а). Дальше вверх по течению, следуя изгибу ледника вглубь суши, поверхность опускалась медленнее, на 20-50 см в год. Кроме того, выше по течению от линии заземления одного из более крупных и быстро текущих притоков ледников (точка B, рис. 1a) поверхность за тот же период понизилась на 30-60 см в год.Надежность этих чисел …
Кроме того, выше по течению от линии заземления одного из более крупных и быстро текущих притоков ледников (точка B, рис. 1a) поверхность за тот же период понизилась на 30-60 см в год.Надежность этих чисел …
Контекст 4
… Методы) выше линии заземления основного быстротекущего ствола ледника (точка A, рис. 1a). Дальше вверх по течению, следуя изгибу ледника вглубь суши, поверхность опускалась медленнее, на 20-50 см в год. Кроме того, выше по течению от линии заземления одного из более крупных и быстро текущих притоков ледников (точка B, рис. 1a) поверхность за тот же период понизилась на 30-60 см в год.Устойчивость этих чисел подтверждается схожестью скоростей изменения высоты по обе стороны от участков с быстрым течением льда от À 10 до þ 20 см в год (рис. 1а). Столь низкие скорости изменения прилегающих участков медленно движущегося льда означают, что колебания в …
Контекст 5
… 1. Кроме того, выше по течению от линии заземления одного из более крупных и быстро текущих притоков ледников (точка B, рис. 1a) поверхность за тот же период понизилась на 30-60 см в год.Устойчивость этих чисел подтверждается схожестью скоростей изменения высоты по обе стороны от участков с быстрым течением льда от À 10 до þ 20 см в год (рис. 1а). Такие низкие темпы изменения в окружающих областях медленно движущегося льда означают, что вариации в локальном поверхностном балансе массы недостаточны для объяснения высоких скоростей опускания поверхности быстро движущегося льда 4. Это убедительно свидетельствует о том, что наблюдаемое истончение основного ствола ледника и притоков связано с динамикой льда….
1a) поверхность за тот же период понизилась на 30-60 см в год.Устойчивость этих чисел подтверждается схожестью скоростей изменения высоты по обе стороны от участков с быстрым течением льда от À 10 до þ 20 см в год (рис. 1а). Такие низкие темпы изменения в окружающих областях медленно движущегося льда означают, что вариации в локальном поверхностном балансе массы недостаточны для объяснения высоких скоростей опускания поверхности быстро движущегося льда 4. Это убедительно свидетельствует о том, что наблюдаемое истончение основного ствола ледника и притоков связано с динамикой льда….
Контекст 6
… насыщенность сигнала и фильтры облачности (см. Методы). Кроме того, в сигнале возвышения поверхности часто преобладает адвекция ярко выраженных топографических особенностей поверхности. Поэтому мы рассматриваем только те области, где поверхность шельфового ледника относительно гладкая, как показывают наши измерения высоты поверхности ICESat-1 и спутниковые снимки (рис. 1a). Соответственно, только сегмент непосредственно после линии заземления вблизи точки C на рис.1а однозначно демонстрирует понижение поверхности на 140-200 см y À 1 …
1a). Соответственно, только сегмент непосредственно после линии заземления вблизи точки C на рис.1а однозначно демонстрирует понижение поверхности на 140-200 см y À 1 …
Контекст 7
… часто преобладает адвекция ярко выраженных топографических особенностей поверхности. Поэтому мы рассматриваем только те области, где поверхность шельфового ледника относительно гладкая, как показывают наши измерения высоты поверхности ICESat-1 и спутниковые снимки (рис. 1a). Соответственно, только на участке, расположенном непосредственно после линии заземления вблизи точки C на рис. 1а, однозначно наблюдается понижение поверхности на 140–200 см / год…
Context 8
… в районе между двумя шельфовыми ледниками вдоль побережья Сабрина, темпы понижения снижаются до B30 см / год (рис. 1а). Вокруг МУГ на большей части площади приземления льда наблюдается небольшое увеличение отметки поверхности, составляющее 0-25 см в год. И снова исключение составляет более быстрый приток ледника (точка D, рис. 1а), который опускается со скоростью B20 см в год. В MUIS сигнал смешанный, на него влияет топография поверхности, как в TIS, с большинством точек…
1а), который опускается со скоростью B20 см в год. В MUIS сигнал смешанный, на него влияет топография поверхности, как в TIS, с большинством точек…
Контекст 9
… в районе между двумя шельфовыми ледниками вдоль побережья Сабрина, темпы понижения снижаются до B30 см г À 1 (рис. 1а). Вокруг МУГ на большей части площади приземления льда наблюдается небольшое увеличение отметки поверхности, составляющее 0-25 см в год. И снова исключение составляет более быстрый приток ледника (точка D, рис. 1а), который опускается со скоростью B20 см в год. В MUIS сигнал смешанный, на него влияет топография поверхности, как и в TIS, при этом большинство точек находится в диапазоне ± 50 см г À 1.Величина декадной поправки на глубину фирна для периода ICESat-1 за период 2003 г.Cat-1, период -2008, в основном составляет от À 30 до þ 10 см год À 1, с площадью MUG …
Контекст 10
.. шельфовый ледник. Наибольшие скорости таяния наблюдаются у восточной окраины обеих полостей под шельфовым ледником, в средней и южной частях TIS и в средней части MUIS (рис. 3). Высокие скорости базального таяния в этих областях могут частично объяснить относительную гладкость соответствующих верхних поверхностей шельфового ледника на спутниковом снимке (рис.1a) Связывание моделированного таяния шельфового ледника и полыньи …
3). Высокие скорости базального таяния в этих областях могут частично объяснить относительную гладкость соответствующих верхних поверхностей шельфового ледника на спутниковом снимке (рис.1a) Связывание моделированного таяния шельфового ледника и полыньи …
Контекст 11
… лучшее соединение TIS, возможно, связано с более длинным фронтом шельфового ледника и более широким входным отверстием вдоль этого фронта. Кроме того, батиметрия перед TIS (дополнительный рис. S1) поддерживает формирование антициклонического круговорота, который переносит теплую воду ко входу в полость TIS. Эти факторы делают полость ТИС более восприимчивой к изменениям условий на континентальном шельфе….
Context 12
… Чтобы проверить, действительно ли утонение TG увеличилось за период ICESat-1, мы подробно исследуем характер изменения высоты поверхности на трех участках (Рис. 1a) в части ледник с наиболее наблюдаемым опусканием поверхности. Профили на рис. 1b показывают, что наибольшее опускание поверхности произошло во второй половине периода (синие точки). Фактически, если рассматривать участки с более медленным течением на флангах ледника (севернее À 67.2 ° и к югу от À 67,8 °) все понижение поверхности …
Профили на рис. 1b показывают, что наибольшее опускание поверхности произошло во второй половине периода (синие точки). Фактически, если рассматривать участки с более медленным течением на флангах ледника (севернее À 67.2 ° и к югу от À 67,8 °) все понижение поверхности …
Контекст 13
… проверить, действительно ли утонение TG увеличилось в течение периода ICESat-1, мы подробно исследуем высоту поверхности закономерности изменения на трех участках (рис. 1а) в части ледника с наиболее наблюдаемым опусканием поверхности. Профили на рис. 1b показывают, что наибольшее опускание поверхности произошло во второй половине периода (синие точки). Фактически, если рассматривать участки с более медленным течением на флангах ледника (севернее À 67.2 ° и южнее À 67,8 °), все понижение поверхности, по-видимому, произошло во второй части периода, поскольку все четыре профиля перекрываются. …
Контекст 14
… еще один В2 см, на основе ошибки В2 миллибара приземного атмосферного давления 43. Таким образом, общая неопределенность возвышений поверхности шельфового ледника составляет B18,5 см. При вычислении изменения отметок поверхности распространение ошибки приводит к неопределенности, вдвое превышающей погрешности отдельных измерений отметки поверхности, как показано на рис.1b и дополнительный рисунок. …
Таким образом, общая неопределенность возвышений поверхности шельфового ледника составляет B18,5 см. При вычислении изменения отметок поверхности распространение ошибки приводит к неопределенности, вдвое превышающей погрешности отдельных измерений отметки поверхности, как показано на рис.1b и дополнительный рисунок. …
Контекст 15
… условия и граничные условия для гидрографии и морского льда предоставляются с грубым разрешением (сетка 18 км), оценивающей циркуляцию и климат океана Фаза II (ECCO2) сопряженный раствор 52. Область модели (дополнительный рис. S1) получена из глобальной конфигурации, используемой ECCO2, но с шагом сетки по горизонтали B1 км и вертикальной дискретизацией 70 вертикальных уровней различной толщины (от 10 м на поверхности до 450 м на самой глубине). уровень)….
РЫБА И ПРОМЫСЛ НА БОЛЬШИХ ВЫСОТАХ: АЗИЯ
РЫБА И ПРОМЫСЛ НА БОЛЬШИХ ВЫСОТАХ: АЗИЯ — ТЕХНИЧЕСКИЙ ДОКУМЕНТ №. 385 по
К. А. Савваитова
А. Савваитова
Биологический факультет МГУ, Воробьевы горы, Москва, 119899, Россия
и
Т. Петр
27 McLeod Street, Toowoomba Qld 4350, Австралия
РЕФЕРАТ
В 1938 году воды озера Севан в Армении начали использоваться для производства гидроэлектроэнергии и для орошения.Самый низкий уровень был достигнут в 1952 году. За этим последовало постепенное повышение уровня воды с возвращением к норме в 1967 году, а затем последующие колебания уровня воды в соответствии с потребностями. В настоящее время уровень воды поддерживается на высоте около 1898 м над уровнем моря, с площадью поверхности 1244 км 2 с ежегодной просадкой для производства гидроэлектроэнергии и орошения. Влияние просадки и колебаний уровня воды на качество воды и рыбные запасы было значительным. До изменения уровня воды в озере в уловах доминировали эндемичная севанская форель ( Salmo ischchan ) и севанская крамуля ( Varicorhinus capoeta) . Забор воды из озера в конечном итоге привел к почти исчезновению севанской форели, которая была внесена в список исчезающих видов рыб. Ухудшение было вызвано двумя основными причинами: высыханием нерестилищ и затопленных макрофитов и эвтрофикацией озера. Управляющие промыслами приняли следующие контрмеры: интродукция пелагического сига ( Coregonus lavaretus ), который в настоящее время составляет 80% от общего улова рыбы, и максимальное минимизация колебаний уровня воды.Планируется постепенно повышать уровень воды, чтобы восстановить некоторые утраченные нерестилища форели, повысить уровень растворенного кислорода и уменьшить эвтрофикацию.
Забор воды из озера в конечном итоге привел к почти исчезновению севанской форели, которая была внесена в список исчезающих видов рыб. Ухудшение было вызвано двумя основными причинами: высыханием нерестилищ и затопленных макрофитов и эвтрофикацией озера. Управляющие промыслами приняли следующие контрмеры: интродукция пелагического сига ( Coregonus lavaretus ), который в настоящее время составляет 80% от общего улова рыбы, и максимальное минимизация колебаний уровня воды.Планируется постепенно повышать уровень воды, чтобы восстановить некоторые утраченные нерестилища форели, повысить уровень растворенного кислорода и уменьшить эвтрофикацию.
1. ВВЕДЕНИЕ
Озеро Севан в Армении (Рис.1) является примером сильно управляемого озера с холодной водой, основной целью которого является производство гидроэлектроэнергии и, в меньшей степени, отвод воды для орошения. Первоначально уровень воды в озере находился на высоте 1906 м над уровнем моря, а площадь озера составляла 1416 км, 2 . Под воздействием сбросов воды на электростанцию и на орошение уровень воды в озере подвергался значительным колебаниям. Сразу после Второй мировой войны уровень воды начал быстро снижаться, и потребовалось 15 лет, прежде чем озеро снова достигло своего полного объема в 1967 году. С тех пор скорость колебаний была менее резкой. В 1990 году уровень озера составлял 1897,5 м, что примерно на 18,5 м ниже номинального уровня воды, и занимало площадь 1244 км 2 (Парпаров, 1990).
Под воздействием сбросов воды на электростанцию и на орошение уровень воды в озере подвергался значительным колебаниям. Сразу после Второй мировой войны уровень воды начал быстро снижаться, и потребовалось 15 лет, прежде чем озеро снова достигло своего полного объема в 1967 году. С тех пор скорость колебаний была менее резкой. В 1990 году уровень озера составлял 1897,5 м, что примерно на 18,5 м ниже номинального уровня воды, и занимало площадь 1244 км 2 (Парпаров, 1990).
Озеро Севан и его ихтиофауна столкнулись с проблемами, так как несколько плотин перекрыли реку Раздан, единственный сток из озера. Обоснование строительства плотины
должен был создать ирригационную и энергетическую систему плотин, которая увеличит скорость сброса из озера, в то же время уменьшив испарение в результате
уменьшение площади озера.Слив воды в озеро, последующие подъемы и колебания уровня привели к необратимому повреждению нерестилищ эндемичной форели, что в сочетании с эвтрофикацией (Оганесян, Парпаров, 1983; Баграмян, 1984) привело к серьезным изменениям в озере. видовой состав рыб. Чтобы остановить дальнейшее ухудшение ситуации и предотвратить дальнейшее падение уровня воды, но в то же время для удовлетворения потребностей ирригации и выработки электроэнергии, был построен ряд насосных станций (Ереванская, Разданская, Татевская, Армянская) и туннель, прорезанный через горы, перекачивает около 200 млн. м 3 3 воды в озеро с водораздела реки Арпа.Позже был построен еще один тоннель для переброски части вод реки Воротан в озеро. Эти меры приносили в общей сложности 420 млн. М3 3 воды в год из новых источников, и ожидается, что в течение первого десятилетия 21 -го века уровень воды в Севане будет повышен на 6 м, т. Е. в свою очередь, изменение эвтрофированного озера обратно к олиготрофному (Баграмян, 1984). Однако может быть слишком поздно для севанской форели, бывшего основного вида рыб, вернуться в том же количестве, что и ранее, что поддерживало прибыльный промысел (Meybeck et al., 1997).
видовой состав рыб. Чтобы остановить дальнейшее ухудшение ситуации и предотвратить дальнейшее падение уровня воды, но в то же время для удовлетворения потребностей ирригации и выработки электроэнергии, был построен ряд насосных станций (Ереванская, Разданская, Татевская, Армянская) и туннель, прорезанный через горы, перекачивает около 200 млн. м 3 3 воды в озеро с водораздела реки Арпа.Позже был построен еще один тоннель для переброски части вод реки Воротан в озеро. Эти меры приносили в общей сложности 420 млн. М3 3 воды в год из новых источников, и ожидается, что в течение первого десятилетия 21 -го века уровень воды в Севане будет повышен на 6 м, т. Е. в свою очередь, изменение эвтрофированного озера обратно к олиготрофному (Баграмян, 1984). Однако может быть слишком поздно для севанской форели, бывшего основного вида рыб, вернуться в том же количестве, что и ранее, что поддерживало прибыльный промысел (Meybeck et al., 1997).
В настоящем документе рассматриваются изменения в рыбных запасах озера Севан в ретроспективе, уделяя особое внимание мерам управления рыболовством, предпринятым в течение этого столетия для противодействия негативным воздействиям, возникающим в результате многократного использования водных ресурсов, где рыболовству уделялось мало внимания. Обсуждается также будущее озерного рыболовства.
Обсуждается также будущее озерного рыболовства.
2. МОРФОМЕТРИЯ И ЛИМНОЛОГИЧЕСКИЕ ХАРАКТЕРИСТИКИ ОЗЕРА СЕВАН
2.1 Морфометрия
Озеро Севан — большое высокогорное пресноводное озеро.Он расположен в северо-восточной части Армении на высоте около 1900 м, в зависимости от водоподачи. Озеро имеет тектоническое происхождение. Его преимущественно треугольная форма имеет удлинение в юго-западном и северо-западном направлениях. Длина озера 75 км, средняя ширина 19 км. Снижение уровня воды на 18,5 м привело к уменьшению площади озера на 12,2%, которая в 1980-е гг. Составляла 1244 км 2 (Парпаров, 1990). Полуостров Артаниш и мыс Норадуз делят озеро на две части: Малый Севан и Большой Севан (рис.2). В озеро впадает 28 малых рек, но из них только один — Раздан. Весенние паводки и таяние снегов — главные источники воды. Сила ветра является важным фактором, влияющим на перемешивание воды, при этом преобладают северные и северо-западные ветры. Если в более глубокой зоне ветер перемешивает только верхние слои воды, то на мелководье — вниз. Дно — песок или песок и глина. Вода озера Севан начала использоваться для выработки электроэнергии в 1938 году, и с тех пор объем воды в озере сократился на 42.2%. Максимальная глубина снижена с 98,7 до 42,2 м на Малом Севане и с 58,7 до 40,2 м на Большом Севане, а средняя глубина с 50,9 до 39,2 м и с 37,7 до 23,6 м соответственно.
Если в более глубокой зоне ветер перемешивает только верхние слои воды, то на мелководье — вниз. Дно — песок или песок и глина. Вода озера Севан начала использоваться для выработки электроэнергии в 1938 году, и с тех пор объем воды в озере сократился на 42.2%. Максимальная глубина снижена с 98,7 до 42,2 м на Малом Севане и с 58,7 до 40,2 м на Большом Севане, а средняя глубина с 50,9 до 39,2 м и с 37,7 до 23,6 м соответственно.
2.2 Температура
Озеро Севан, расположенное на широте 40 o северной широты, является довольно теплым озером. Среднегодовая температура поверхностных вод составляет 10-12 o C. Минимальная температура воды 1-2 o C достигается в период с января по апрель.Зимой вода не расслаивается. Озеро замерзает каждый год. В августе максимальная температура поверхностной воды может достигать 19 o С. Летом вода стратифицирована, гиполимнион находится на глубине 25-30 м. С октября по декабрь водная толща озера перемешивается сверху вниз, с одинаковой температурой повсюду (Гезалян, 1979).
Температура воды начинает повышаться в первой половине марта. У устьев рек, впадающих в озеро, вода прогревается быстрее, чем в основной части озера.Термическая стратификация воды в верхних 10 м начинает развиваться в конце мая. В июне эпилимнион на Малом Севане уже достигает 15-20 м, что близко к эпилимниону Большого Севана. В июне или июле в озере развиваются горизонтальные градиенты температуры. Вода начинает остывать в сентябре и продолжается до первой половины января. Перемешивание воды под действием ветра способствует нарушению стратификации. В Большом Севане вода полностью перемешивается в октябре-ноябре при температуре воды 11-12 o C; в Малом Севане это происходит при температуре 5 o C в ноябре-декабре.В настоящее время вода в Большом Севане начинает перемешиваться на месяц раньше и имеет температуру на 3-4 o C выше, чем в прошлом, до манипуляций с уровнем воды. В Малом Севане понижение уровня воды не повлияло на возникновение перемешивания вод и термической стратификации. С понижением уровня воды осенняя температура воды у дна повышается. До техногенного падения уровня воды температура воды в гиполимнионе Большой Севан не превышала 8 o ° C.С 1965 года температура достигла почти 15 o C.
С понижением уровня воды осенняя температура воды у дна повышается. До техногенного падения уровня воды температура воды в гиполимнионе Большой Севан не превышала 8 o ° C.С 1965 года температура достигла почти 15 o C.
Раньше озеро замерзало раз в 3 или 4 года, но теперь оно замерзает ежегодно, и ледяной покров, толщина которого может достигать 30-40 см, может оставаться с конца января до начала апреля. Подо льдом вода может иметь обратную стратификацию, при этом верхние слои имеют температуру, близкую к нулю, а нижний слой 1,2–2,3 o ° C (Гезалян, 1979).
2.3 Концентрация растворенного кислорода
Концентрация растворенного кислорода в литоральной зоне находится в пределах 6.23 и 11,08 мг л -1 , а в центральной части озера между 4,56 и 11,6 мг л -1 (Гезалян, Хорлашко, 1979). В 1928 г. наибольшее содержание растворенного кислорода было зарегистрировано весной, после тщательного перемешивания толщи воды, при температуре воды 4 o С. Осенью концентрация растворенного кислорода снижалась. Средний минимум кислорода никогда не был ниже 5,6 мг / л -1 , максимум достигал 11,8 мг / л -1 (Лятти, 1932). В 1947 году самая низкая концентрация кислорода была достигнута в ноябре.С 1970-х годов концентрация растворенного кислорода у дна стала очень низкой по всему озеру с нулевыми значениями, достигнутыми в конце периода стратификации (Парпаров, 1990). Эта ситуация, по-видимому, напрямую связана с эвтрофикацией.
Осенью концентрация растворенного кислорода снижалась. Средний минимум кислорода никогда не был ниже 5,6 мг / л -1 , максимум достигал 11,8 мг / л -1 (Лятти, 1932). В 1947 году самая низкая концентрация кислорода была достигнута в ноябре.С 1970-х годов концентрация растворенного кислорода у дна стала очень низкой по всему озеру с нулевыми значениями, достигнутыми в конце периода стратификации (Парпаров, 1990). Эта ситуация, по-видимому, напрямую связана с эвтрофикацией.
2,4 Химический состав воды
Сравнение химического состава воды (Таблица 1) показывает изменения с 1929 по 1983 год. Озеро имело высокий pH 8,4. Среди доминирующих катионов не было изменений концентрации Mg и умеренных изменений концентраций Ca, Na и K.Преобладали бикарбонаты. Концентрации сульфатов не изменились, но было небольшое увеличение хлоридов. Данные не свидетельствуют об изменении концентрации Si. Произошло заметное снижение растворенного кислорода, при этом в последние годы было зафиксировано полное обескислороживание воды. Это также объясняет увеличение содержания нитратного / нитритного азота, аммиака и железа (не в таблице 1), где Парпарова (1979) обнаружила 0,014 мг л -1 .
Это также объясняет увеличение содержания нитратного / нитритного азота, аммиака и железа (не в таблице 1), где Парпарова (1979) обнаружила 0,014 мг л -1 .
Изменения растворенного кислорода и увеличение содержания азота и железа являются индикаторами процесса эвтрофикации в озере Севан.Основные причинные факторы, лежащие в основе этого, видны в постепенном понижении уровня воды, которое сделало озеро более мелким, и в увеличении сброса в озеро плохо очищенных сточных вод.
2,5 Гидробиологические характеристики
С 1960-х годов в озере происходят изменения в составе фитопланктона, зоопланктона и бентоса. До начала необратимого осушения озера в пелагиали озера числилось 26 видов водорослей.Наиболее распространенными из них были диатомеи Asterionella formosa , Stephanodiscus astraea и S. hantzschii , зеленые водоросли Gloeococcus schroeteri и род Oocystis. В то время количество видов было невелико. С 1964 года бурный рост сине-зеленых водорослей, таких как Anabaena lemmermanii , стал обычным явлением в озере, вызвав цветение (Легович, 1979). В 1970-х годах состав фитопланктона изменился: появились новые виды, а некоторые из прежних видов исчезли.Большинство новых видов водорослей принадлежало к Protococcales, в том числе Ankistrodesmus angustus, Hyaloraphidium rectum , Lagerheimia , Golenkiniopsis , Treubaria, Tetrastrum и некоторые другие. Род Euglena прижился в пелагиали. Диатомовые водоросли Melosira italica , Peridinium sp., Closterium и Mougeotia sp. появились (Казарян, 1979). Сравнение ситуации 1952 г. и 1979 г. показало, что биомасса фитопланктона увеличилась в 22 раза, т.е.е. до 18 г м -3 в Большом Севане и в 10 раз, т.е. до 7,3 г м -3 в Малом Севане. Произошло значительное увеличение биомассы Melosira italica до 3 г -3 , что соответствует 15 миллионам особей на литр, и Hyaloraphidium rectum (0,32 г -3 , 61 миллион экз.
С 1964 года бурный рост сине-зеленых водорослей, таких как Anabaena lemmermanii , стал обычным явлением в озере, вызвав цветение (Легович, 1979). В 1970-х годах состав фитопланктона изменился: появились новые виды, а некоторые из прежних видов исчезли.Большинство новых видов водорослей принадлежало к Protococcales, в том числе Ankistrodesmus angustus, Hyaloraphidium rectum , Lagerheimia , Golenkiniopsis , Treubaria, Tetrastrum и некоторые другие. Род Euglena прижился в пелагиали. Диатомовые водоросли Melosira italica , Peridinium sp., Closterium и Mougeotia sp. появились (Казарян, 1979). Сравнение ситуации 1952 г. и 1979 г. показало, что биомасса фитопланктона увеличилась в 22 раза, т.е.е. до 18 г м -3 в Большом Севане и в 10 раз, т.е. до 7,3 г м -3 в Малом Севане. Произошло значительное увеличение биомассы Melosira italica до 3 г -3 , что соответствует 15 миллионам особей на литр, и Hyaloraphidium rectum (0,32 г -3 , 61 миллион экз.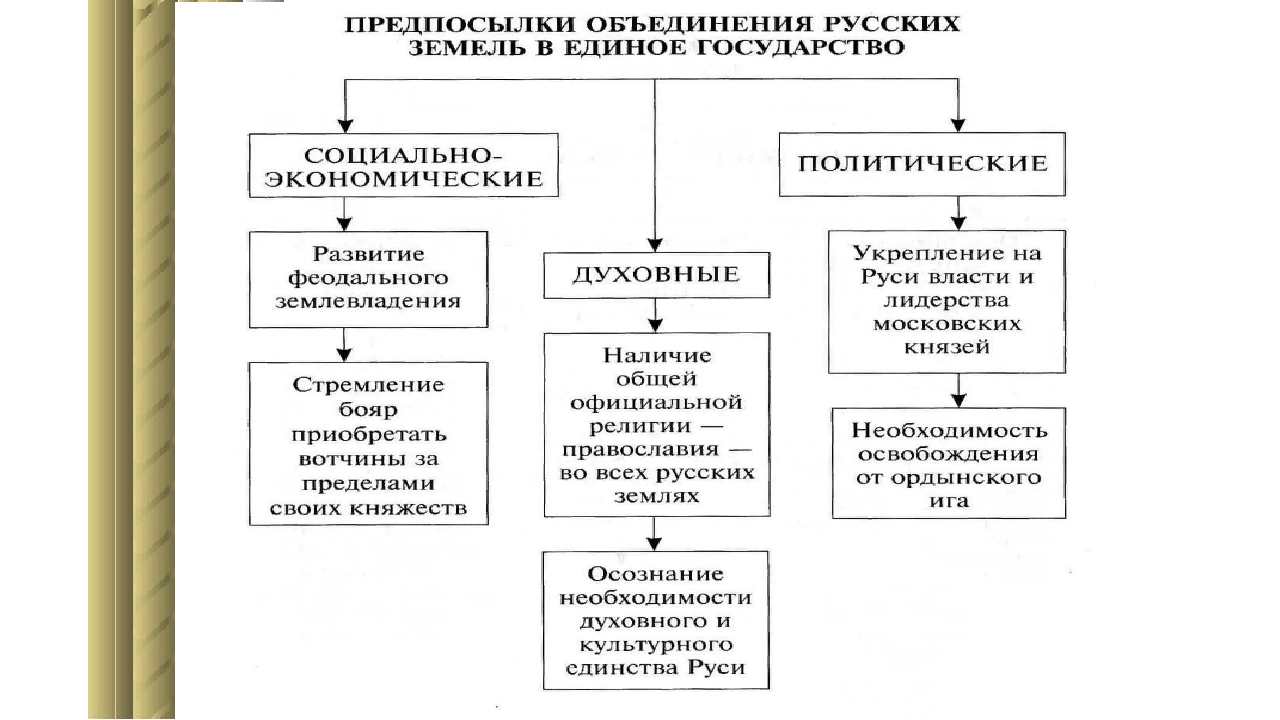 Л -1 ) (Казарян, 1979).
Л -1 ) (Казарян, 1979).
До падения уровня воды в 1938 году в флоре дна озера до глубины 14 м преобладали каменистый ( Chara ) и мох.Но после этого количество видов и их биомасса упали до одной десятой от предыдущего уровня. К 1976 г. практически исчезли Chara и ранее распространенные Potamogeton natans, P. pusillus, P. densus и Zannichelia palustris (Гамбарян, 1979).
Когда в 1938 году уровень воды начал падать, состав зоопланктона оставался стабильным в течение длительного времени. Всего было 10 видов коловраток, кладоцер и веслоногих рачков.Самая высокая биомасса была у веслоногих ракообразных, которые составляли около 60% от общего количества, и у кладоцера Daphnia (Таблицы 2 и 3). Постепенно видовой состав стал меняться, что сопровождалось изменением биомассы отдельных видов. В 1974-1980 гг. Доминировал зоопланктон меньшего размера, при этом на коловратки приходилось до 80% биомассы. Ситуация снова изменилась в течение 1981-1985 годов, когда биомасса коловраток снизилась до 60-65% от общей биомассы, а биомасса дафний Daphnia и Cyclops увеличилась (Симонян, 1988). Таким образом, можно выделить три основных периода биомассы зоопланктона: 1937-1969 гг. — низкая биомасса; 1972-1979 — высокая биомасса; 1980-1984 гг. — низкая биомасса (табл. 4).
Таким образом, можно выделить три основных периода биомассы зоопланктона: 1937-1969 гг. — низкая биомасса; 1972-1979 — высокая биомасса; 1980-1984 гг. — низкая биомасса (табл. 4).
В зообентосе озера Севан преобладают личинки хирономид ( Chironomus plumosus, C. markosjani, Procladius sp., Glyptotendipes barbipes ), Oligochaeta ( Potamothrix hammoniensis, Tubifedeulsella , Herpolusa et al. Glossiphonia Complanata ), двустворчатые моллюски ( Pisidium nitidus и P.casertanum ), улитки ( Valvata piscinalis, Limnaea stagnalis, Radix ovata ), поденки (представлены в озере только одним видом Caenis modesta ) и гаммариды ( Gammarus lacustris) (Arnoldi, 1983; Arnoldi, 1929; ). В 1929 г. в бентосе преобладали гаммариды и хирономиды, остальные группы составляли лишь незначительную часть биомассы. Это было типичным представителем олиготрофного характера Севана в том году. За 1938-1948 гг. Олигохет представляли 36 человек. 2% биомассы, гаммариды 34,6% (Фридман, 1950). В 1955-1967 гг. По-прежнему доминировали олигохеты, за ними следовали хирономиды, при этом биомасса гаммарид снизилась, а биомасса моллюсков увеличилась более чем вдвое. Пиявки остались неизменными (Маркосян, 1959; Мешкова, 1976). Совсем недавно биомасса хирономид повысилась до исходного уровня (60,5%), а биомасса олигохет — до 37,7%; биомасса моллюсков увеличилась в три раза по сравнению с периодом 1938-1948 гг. (табл. 5). Тем не мение. Биомасса гаммарид резко снизилась в результате сокращения площади и биомассы подводных водных макрофитов, а также площади каменистого и песчаного дна (Маркосян, 1959).Увеличивается также добыча гаммарид сигами-сигами (Дадикян, 1955; Пивазян, 1984).
2% биомассы, гаммариды 34,6% (Фридман, 1950). В 1955-1967 гг. По-прежнему доминировали олигохеты, за ними следовали хирономиды, при этом биомасса гаммарид снизилась, а биомасса моллюсков увеличилась более чем вдвое. Пиявки остались неизменными (Маркосян, 1959; Мешкова, 1976). Совсем недавно биомасса хирономид повысилась до исходного уровня (60,5%), а биомасса олигохет — до 37,7%; биомасса моллюсков увеличилась в три раза по сравнению с периодом 1938-1948 гг. (табл. 5). Тем не мение. Биомасса гаммарид резко снизилась в результате сокращения площади и биомассы подводных водных макрофитов, а также площади каменистого и песчаного дна (Маркосян, 1959).Увеличивается также добыча гаммарид сигами-сигами (Дадикян, 1955; Пивазян, 1984).
3. РЫБНАЯ ФАУНА ОЗЕРА СЕВАН
В озере Севан обитает эндемичная ихтиофауна, представленная всего тремя видами: севанская форель ( Salmo ischchan Kessler), севанская крамуля ( Varicorhinus capoeta sevangi Filippi) и севанский усач ( Barbus lacerta goktchaessicus Kessler). Однако в прошлом для увеличения уловов было завезено больше видов рыб: ладожский сиг ( Coregonus lavaretus ladoga ) из Ладожского озера и сиг из Чудского озера ( Coregonus lavaretus maraenoides ).Оба вида, интродуцированные в 1924 году, не только прижились на Севане, но и скрестились, дав новый гибрид Coregonus lavaretus sevanicus Dadikjan (Дадикян, 1986).
Однако в прошлом для увеличения уловов было завезено больше видов рыб: ладожский сиг ( Coregonus lavaretus ladoga ) из Ладожского озера и сиг из Чудского озера ( Coregonus lavaretus maraenoides ).Оба вида, интродуцированные в 1924 году, не только прижились на Севане, но и скрестились, дав новый гибрид Coregonus lavaretus sevanicus Dadikjan (Дадикян, 1986).
Карп гибель ( Carassius auratus gibelio Bloch) также был случайно завезен в начале 1980-х годов и создал сильные запасы в районах, богатых водными макрофитами.
Хотя севанская форель и севанская храмуля были обычным явлением в озере до того, как уровень воды начал снижаться, их запасы позже резко сократились.В 1984 г. оба вида занесены в Красную книгу СССР (1984 г.). Запасы севанского усачка также сократились, и этот вид занесен в Красную книгу Армении. Запасы Храмуля остаются стабильными, а запасы сиговых рыб сильно увеличиваются.
3.1 Биология севанских рыб
3. 1.1 Севанская форель — Салмо ищчан
1.1 Севанская форель — Салмо ищчан
Севанская форель представлена четырьмя подвидами, отличающимися друг от друга по ряду морфологических признаков.
« Зимняя» Севанская форель ( Салмо ищчан ищчан )
В прошлом озеро было обильно заселено так называемой «зимней» севанской форелью, строго озерной формой, ищущей пищу и нерестящейся нигде, кроме озера. Этот подвид делится на два типа: альфа и бета (Фортунатов, 1927). Альфа-тип нерестится с начала ноября до конца декабря на мелководье (глубина 0,5-5 м) на песке и гравии литорали вдоль северо-западного берега озера. .Нерестилища этого типа занимали 17 га (Павлов, 1951). Бета-тип нерестился с января по март в юго-восточной части Большого Севана, на глубине до 22 м. Нерестилища данного типа занимали 5 га. Альфа-тип нерестился при температуре 1-3 o ° C, бета-тип при -0,3-3 ° ° C (Фортунатов и др. , 1932; Дадикян, 1986). Альфа достигла половой зрелости в возрасте 5-6 лет, бета — через два года и с тех пор нерестилась ежегодно (Дадикян, 1986). В отличие от других подвидов, «зимняя» севанская форель была очень плодовита, давая до 8700 яиц (Павлов, 1951).Рыба вырастала до 90 см в длину и до 15 кг и быстро выращивала. Мертвых особей длиной 80 см и весом около 8 кг часто видели в озере до того, как уровень воды начал падать. «Зимняя» севанская форель питалась бентосом: Gammmarus составляла 90-95% от общего веса корма (Дадикян, 1986). Озеро давало ежегодно около 150 тонн «зимней» севанской форели. Альфа-тип был более распространен, чем бета-тип. Однако с понижением уровня воды альфа-тип потерял свои нерестилища, а когда падение уровня воды достигло 10 м, этот тип вымер.Бета-тип выжил, потому что его нерестилища были расположены глубже и, следовательно, не подвергались воздействию. Однако его уловы стали незначительными (Дадикян, 1986).
В отличие от других подвидов, «зимняя» севанская форель была очень плодовита, давая до 8700 яиц (Павлов, 1951).Рыба вырастала до 90 см в длину и до 15 кг и быстро выращивала. Мертвых особей длиной 80 см и весом около 8 кг часто видели в озере до того, как уровень воды начал падать. «Зимняя» севанская форель питалась бентосом: Gammmarus составляла 90-95% от общего веса корма (Дадикян, 1986). Озеро давало ежегодно около 150 тонн «зимней» севанской форели. Альфа-тип был более распространен, чем бета-тип. Однако с понижением уровня воды альфа-тип потерял свои нерестилища, а когда падение уровня воды достигло 10 м, этот тип вымер.Бета-тип выжил, потому что его нерестилища были расположены глубже и, следовательно, не подвергались воздействию. Однако его уловы стали незначительными (Дадикян, 1986).
Гегаркуни Севанская форель ( Салмо ищчан гегаркуни )
Раньше севанская форель гегаркуни была обыкновенной севанской рыбой. В настоящее время он присутствует только в Большом Севане. Зимой держится на открытых участках озера. С апреля она выходит на литораль, а летом, по мере прогрева воды, рыба попадает в более глубокие воды (Пивазян, 1979).Фортунатов (1927) описал два вида севанской форели гегаркуни: альфа и бета. Альфа выросла до средней длины 40 см и нерестилась с сентября по январь в реках Гаварагет, Цаккар, Макенис и Масрик. Бета-версия была тяжелее, доходила до 4 кг. С мая по июнь мигрировал из Севана в реки Аргичи и Гезал-Дара, в ручьи Бахтак и Балык, где осенью нерестился в верховьях на высоте более 2500 м. Бета-версии больше не существует; альфа-тип, неспособный к естественному воспроизводству, искусственно разводится в инкубаториях.Форель Гегаркуни Севана достигает половой зрелости в возрасте 3-6 лет. Повторный нерест случается редко. С началом снижения уровня воды запасы этой форели помолодели, рост и плодовитость снизились, и теперь самцы преобладают над самками (Таблица 6).
Зимой держится на открытых участках озера. С апреля она выходит на литораль, а летом, по мере прогрева воды, рыба попадает в более глубокие воды (Пивазян, 1979).Фортунатов (1927) описал два вида севанской форели гегаркуни: альфа и бета. Альфа выросла до средней длины 40 см и нерестилась с сентября по январь в реках Гаварагет, Цаккар, Макенис и Масрик. Бета-версия была тяжелее, доходила до 4 кг. С мая по июнь мигрировал из Севана в реки Аргичи и Гезал-Дара, в ручьи Бахтак и Балык, где осенью нерестился в верховьях на высоте более 2500 м. Бета-версии больше не существует; альфа-тип, неспособный к естественному воспроизводству, искусственно разводится в инкубаториях.Форель Гегаркуни Севана достигает половой зрелости в возрасте 3-6 лет. Повторный нерест случается редко. С началом снижения уровня воды запасы этой форели помолодели, рост и плодовитость снизились, и теперь самцы преобладают над самками (Таблица 6).
« Лето» Севанская форель ( Salmo ischchan aestivalis )
«Летняя» севанская форель встречается как в озерной, так и в речной форме. В прошлом он присутствовал в юго-западной, юго-восточной и северо-западной частях озера (Павлов, 1951).Позже он был обнаружен по всему озеру, но в конце 1980-х годов в основном ограничивался Большим Севаном (Савваитова и др. , 1989; Павлов и др. , 1989). При полном уровне воды рыба нерестилась в реках и в озере (Фортунатов, 1927) на гравии. С мая по июль строил гнезда в более медленных частях рек (Лещинская, 1950). Ее биологические характеристики и изменения, произошедшие за период понижения уровня воды, приведены в Таблице 6. В настоящее время «летняя» севанская форель мигрирует на нерест в реки Макенис, Масрик, Личк и Цаккар, которые перекрыты плотинами для обеспечения возможности отлов маточного стада; собирают икру и возвращают рыбу в озеро.
В прошлом он присутствовал в юго-западной, юго-восточной и северо-западной частях озера (Павлов, 1951).Позже он был обнаружен по всему озеру, но в конце 1980-х годов в основном ограничивался Большим Севаном (Савваитова и др. , 1989; Павлов и др. , 1989). При полном уровне воды рыба нерестилась в реках и в озере (Фортунатов, 1927) на гравии. С мая по июль строил гнезда в более медленных частях рек (Лещинская, 1950). Ее биологические характеристики и изменения, произошедшие за период понижения уровня воды, приведены в Таблице 6. В настоящее время «летняя» севанская форель мигрирует на нерест в реки Макенис, Масрик, Личк и Цаккар, которые перекрыты плотинами для обеспечения возможности отлов маточного стада; собирают икру и возвращают рыбу в озеро.
Боджак Севанская форель ( Salmo ischchan danilewskii )
Карликовая, медленнорастущая озерная рыба (таблица 6). Отличительная особенность рыбы в том, что она не мигрирует. Его места нагула и нерестилища совпадают. При стабильном уровне воды в озере этот подвид населял в основном Малый Севан и западные прибрежные районы Большого Севана. Нерестится в октябре-ноябре на глубине 0,5-15 м при температуре 10–90–19–90–120 ° С (Фортунатов, 1927; Павлов, 1951).После того, как в 1963-1965 годах нерестилища стали постоянно обнажаться и высыхать, рыба переместилась на новые нерестилища, время нереста изменилось, и она нерестилась в более прохладной воде, чем раньше.
При стабильном уровне воды в озере этот подвид населял в основном Малый Севан и западные прибрежные районы Большого Севана. Нерестится в октябре-ноябре на глубине 0,5-15 м при температуре 10–90–19–90–120 ° С (Фортунатов, 1927; Павлов, 1951).После того, как в 1963-1965 годах нерестилища стали постоянно обнажаться и высыхать, рыба переместилась на новые нерестилища, время нереста изменилось, и она нерестилась в более прохладной воде, чем раньше.
3.1.2 Севан храмуля — Varicorhinus capoeta sevangi
Севанская храмуля имеет изолированные популяции, разбросанные по всему озеру. Речные популяции присутствуют в более крупных реках, впадающих в озеро (Дадикян, 1986). Период нереста в озере и реках длится с мая по август.Нерест начинается при температуре 12 o ° C и достигает пика при температуре воды 15 o ° C в июне-июле. Храмуля нерестится на песке и гравии, свободном от глины и водных растений. С падением уровня воды рыба потеряла большую часть своих нерестилищ, и это увеличило важность речных нерестилищ, особенно в реках Варденис и Аргичи. Если раньше при полном уровне воды в озере самцы этого вида достигали половой зрелости в возрасте 6+ лет, а самки — в 11+ лет, то сегодня они созревают раньше.Рыба может достигать возраста 24 лет, но обычный возраст естественной смертности составляет 14-16 лет. С падением уровня воды запасы севанской храмули помолодели, и скорость воспроизводства существенно изменилась (Габриелян, Тряпицына, 1984). С понижением верхнего предела возраста плодовитость заметно снизилась (Габриелян, Шатуновский, 1984), теперь она составляет от 23 000 до 75 000 яиц. Храмуля питается водными макрофитами, а с возраста 2+ еще и детритом (Дадикян, 1986). С 1936 г., в связи с постепенным ухудшением условий воспроизводства из-за постоянного воздействия на многие нерестилища и утратой многих нерестилищ в реках, запасы крамули в Севане постепенно сокращаются (Габриелян и др., 1990), хотя все еще поддерживается достаточно здоровый уровень для поддержания ограниченного промысла.
Если раньше при полном уровне воды в озере самцы этого вида достигали половой зрелости в возрасте 6+ лет, а самки — в 11+ лет, то сегодня они созревают раньше.Рыба может достигать возраста 24 лет, но обычный возраст естественной смертности составляет 14-16 лет. С падением уровня воды запасы севанской храмули помолодели, и скорость воспроизводства существенно изменилась (Габриелян, Тряпицына, 1984). С понижением верхнего предела возраста плодовитость заметно снизилась (Габриелян, Шатуновский, 1984), теперь она составляет от 23 000 до 75 000 яиц. Храмуля питается водными макрофитами, а с возраста 2+ еще и детритом (Дадикян, 1986). С 1936 г., в связи с постепенным ухудшением условий воспроизводства из-за постоянного воздействия на многие нерестилища и утратой многих нерестилищ в реках, запасы крамули в Севане постепенно сокращаются (Габриелян и др., 1990), хотя все еще поддерживается достаточно здоровый уровень для поддержания ограниченного промысла.
3. 1.3 Севанский усач — Barbus lacerta goktchaicus
1.3 Севанский усач — Barbus lacerta goktchaicus
В системе озера Севан / реки существуют три формы этой рыбы: озерная форма, переходная форма озера-реки и речная форма. Форма озера строго ограничена озером, где обитает травертин и каменистое дно. Он вырастает до максимального размера 30 см и веса 500 г, а некоторые особи достигают возраста 12+.Это медленный производитель. До того, как уровень воды начал снижаться, озерная форма севанского усачей была самой распространенной из трех форм, давая урожайность до 12 тонн в год. Снижение уровня воды и потеря многих нерестилищ привели к резкому сокращению рыбных запасов этой формы. Переходная форма озеро-река большую часть времени проводит в озере, но нерестится в реках. Улавливается около 4 т в год, часть — в реках. Такой промысел может привести к полному удалению всех производителей. Река является самой маленькой из трех, она ограничена средним и верхним течениями рек на высоте около 2200 м.
Озёрная форма созревает в возрасте 4-6 лет; река сформировалась годом ранее. Нерест начинается в июне при температуре воды около 14 o C и заканчивается в августе при температуре 18 o C. Откладывают две партии яиц с интервалом в 12-15 дней. Мальки рыб питаются зоопланктоном; молодь питается в основном гаммаридами и ручейниками, а также икрой форели в период нереста форели. Частично речная форма питается и летающими насекомыми (Дадикян, 1986).
Нерест начинается в июне при температуре воды около 14 o C и заканчивается в августе при температуре 18 o C. Откладывают две партии яиц с интервалом в 12-15 дней. Мальки рыб питаются зоопланктоном; молодь питается в основном гаммаридами и ручейниками, а также икрой форели в период нереста форели. Частично речная форма питается и летающими насекомыми (Дадикян, 1986).
Севанский усач интенсивно питается с мая по октябрь.В холодное время года рыба мало питается, оставаясь под камнями, в основном, неподвижно, до весны.
Севанский усач — рыба не обычная. В прошлом это составляло менее 3% от общего улова. С тех пор это стало редкостью.
3.1.4 Севанский сиг — Coregonus lavaretus sevanicus
Два подвида сига были завезены на Севан в 1924-1927 гг .: ладожский сиг ( C.л. ладога ) и Чудский сиг ( C. l. maraenoides ). Позже они создали гибрид. Теперь рыба размножается в озере.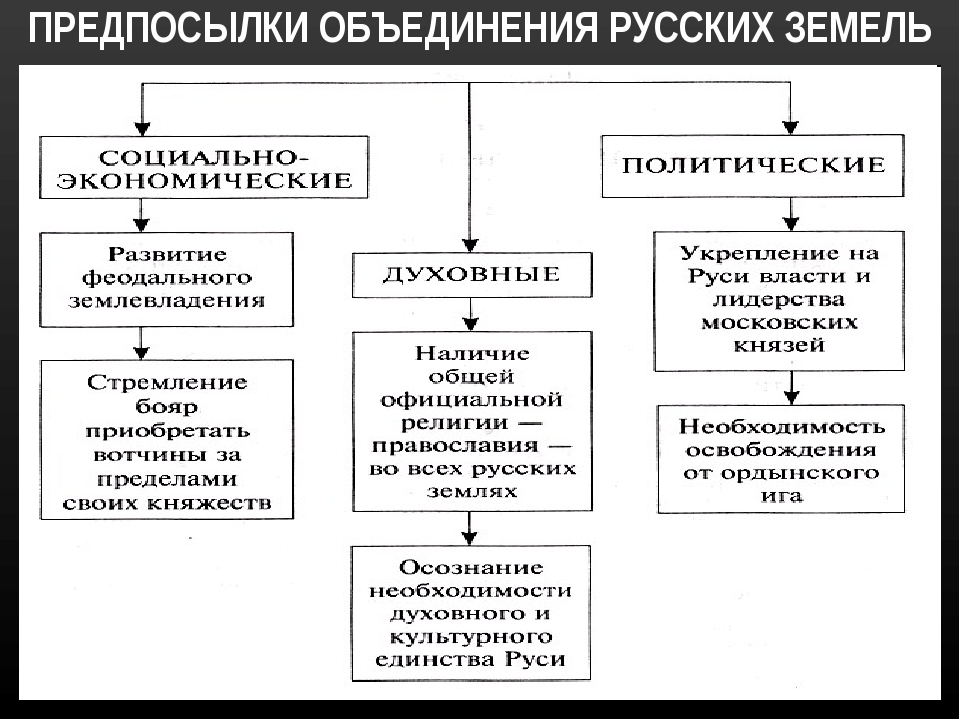 Они нерестятся на литорали на глубине 0,5-10 м, в основном в юго-восточной части озера (Дадикян, 1986; Смолей, и др., , 1985). После их интродукции рыба начала нереститься с ноября по январь. Средняя длина тела расплода составляла от 35 до 50 см, плодовитость от 20 000 до 31 400 особей. Завезенные рыбы росли быстрее и созревали раньше на Севане, чем в первоначальном ареале (Южакова, 1985).В 1940-1950-х годах севанский сиг нерестился в возрасте 9-10 лет и старше (Павлов, 1947; Маилян, 1957), но в уловах преобладали 3-4-летние возрастные группы. Постепенно возрастная структура сига изменилась, и поголовье стало значительно моложе (Южакова, 1979): резко увеличилась доля рыб, достигающих половой зрелости в возрасте от 1 года до 2 лет (длина тела 28-37 см). . Это было замечено в результате повышения температуры воды в озере по мере того, как озеро становилось все более мелким.В центральной части озера температура повысилась на 4-5 o C, а на литорали с 10 o C до 16-18 o C.
Они нерестятся на литорали на глубине 0,5-10 м, в основном в юго-восточной части озера (Дадикян, 1986; Смолей, и др., , 1985). После их интродукции рыба начала нереститься с ноября по январь. Средняя длина тела расплода составляла от 35 до 50 см, плодовитость от 20 000 до 31 400 особей. Завезенные рыбы росли быстрее и созревали раньше на Севане, чем в первоначальном ареале (Южакова, 1985).В 1940-1950-х годах севанский сиг нерестился в возрасте 9-10 лет и старше (Павлов, 1947; Маилян, 1957), но в уловах преобладали 3-4-летние возрастные группы. Постепенно возрастная структура сига изменилась, и поголовье стало значительно моложе (Южакова, 1979): резко увеличилась доля рыб, достигающих половой зрелости в возрасте от 1 года до 2 лет (длина тела 28-37 см). . Это было замечено в результате повышения температуры воды в озере по мере того, как озеро становилось все более мелким.В центральной части озера температура повысилась на 4-5 o C, а на литорали с 10 o C до 16-18 o C. Также считается, что жизненный цикл этого вида сократился. в результате интенсивной промысловой нагрузки на рыбные запасы (Южакова, 1985).
Также считается, что жизненный цикл этого вида сократился. в результате интенсивной промысловой нагрузки на рыбные запасы (Южакова, 1985).
В то время как первоначальная цель интродукции сига заключалась в обеспечении лучшего использования зоопланктона, у сига двухлетнего и старше гаммариды составляли основной корм.Это привело к конкуренции за корм с севанской форелью (Фортунатов и др. , 1932; Маилян, 1957). Через несколько лет давление питания, а также сокращение количества погруженных макрофитов снизили биомассу гаммарид, и сигу пришлось вернуться к зоопланктону (Пивазян, 1982).
Запасы сига оставались на низком уровне до 1955 года. Падение уровня воды в озере на 16 м и эвтрофикация, особенно мелководья, улучшили условия для сига, и впоследствии их запасы увеличились, причем быстрее всего в начале эвтрофикации.С увеличением количества сига в озере коммерческий промысел сига улучшился, и к концу 1960-х годов сиг стал основной промысловой рыбой. Сегодня они составляют около 80% от общего улова около 1000 т.
4. ИЗМЕНЕНИЯ В ЗАПАСАХ РЫБЫ ПРИ ПОПАДАНИИ УРОВНЯ ВОДЫ ОЗЕРА
4.1 Факторы сокращения рыбных запасов
До начала 1960-х годов озеро было олиготрофным. За этим последовала постепенная эвтрофикация в результате антропогенных воздействий, таких как необратимый слив воды из озера для гидроэнергетики, и увеличение поступления загрязняющих веществ из растущего числа населенных пунктов вокруг озера.Озеро становилось все более мелким, а диапазон температур воды увеличивался, и теперь поверхность озера зимой регулярно покрывалась льдом. Наблюдалась повышенная мутность воды, снижение концентрации растворенного кислорода и периодическая дезоксигенация придонных слоев воды, приводящая к сокращению водных макрофитов, в то время как увеличение сине-зеленых водорослей приводило к «цветению воды». Дно озера также покрылось глиной (табл. 7). Это сопровождалось увеличением биомассы и продукции зоопланктона, придонных хирономид и олигохет.Улов рыбы увеличился с 10,0 до 20,9 кг / га -1 во второй половине 1970-х годов (таблица 7). Произошла резкая перемена в преобладании видов рыб. Запасы севанской форели резко сократились, а интродуцированный сиг увеличился со 166,1 т в 1960 г. до 846,3 т в 1973 г. (Таблица 8). Храмуля сохранила первоначальную плотность (Южакова, Пивазян, 1975). Севанский усач, годовой улов которого до падения уровня воды составлял 20 т, стал редкостью из-за плохих условий для его нереста.
Произошла резкая перемена в преобладании видов рыб. Запасы севанской форели резко сократились, а интродуцированный сиг увеличился со 166,1 т в 1960 г. до 846,3 т в 1973 г. (Таблица 8). Храмуля сохранила первоначальную плотность (Южакова, Пивазян, 1975). Севанский усач, годовой улов которого до падения уровня воды составлял 20 т, стал редкостью из-за плохих условий для его нереста.
Озеро Севан с преобладанием форели (60-65% от общего промыслового улова) когда-то было уникальным водоемом. Популяция форели была высокой из-за благоприятных условий для размножения, поддерживаемых зарыблением, а также из-за обилия гаммарид, являющихся обильным источником пищи для форели. Причем гаммариды не употреблялись в пищу другими рыбами. Падение уровня воды затронуло нерестилища всех подвидов и форм севанской форели, кроме исходной S. i. gegarkuni и S.я. a estivalis , потому что инкубатории были в состоянии производить достаточное количество посадочного материала. Когда уровень озера начал падать, в течение ряда лет большое количество яиц (до 70 миллионов ежегодно) собиралось и инкубировалось в инкубаториях для производства зарыбленного материала для выпуска в озеро. Несмотря на все усилия, запасы севанской форели начали сокращаться из-за новых факторов, которые повлияли на среду обитания форели: увеличение забора воды для орошения и увеличение сброса агрохимикатов в ручьи и реки, стекающие с орошаемых полей, а также попадание промышленных и муниципальных загрязняющих веществ. реки и ручьи, используемые для производства зверобоя.Кроме того, усилилась конкуренция за пищу со стороны интродуцированного сига (Пивазян, 1972, 1979).
Несмотря на все усилия, запасы севанской форели начали сокращаться из-за новых факторов, которые повлияли на среду обитания форели: увеличение забора воды для орошения и увеличение сброса агрохимикатов в ручьи и реки, стекающие с орошаемых полей, а также попадание промышленных и муниципальных загрязняющих веществ. реки и ручьи, используемые для производства зверобоя.Кроме того, усилилась конкуренция за пищу со стороны интродуцированного сига (Пивазян, 1972, 1979).
Браконьерство также несет существенную ответственность за сокращение запасов севанской форели. Это стало настолько серьезным, что угрожает самому существованию вида.
С падением уровня воды нерестилища севанского храмуля исчезли, так как в полном озере они находились всего на 1 м ниже поверхности воды.С 60-х годов прошлого века запасы храмули подверглись перелову.
4.2 Изменение уловов рыбы
До того, как уровень воды в озере Севан начал падать, в уловах преобладала зимняя, летняя и гегаркунинская форель, реже — боджакская форель. В 1970 г. в уловах преобладала гегаркунинская форель — 120 т в год, за ней следовала «летняя» форель. В 1975 г. гегаркунинская форель составляла 56-70% уловов, «летняя» форель — около 20%, а «зимняя» форель и боджак составляли 7-18% (Смолей, 1979, 1979а).Это было связано с разным воздействием падения уровня воды на нерестилища каждого из подвидов. Основные нерестилища «зимней» форели высохли в 1963 году, и аквакультурам не удалось найти подходящий метод инкубации икры «зимней» форели. Хотя нерестилища «летней» форели в озере высохли намного раньше, т. Е. В 1948 году, чем у «зимней» форели, запасы оставались стабильными до 1960-х годов, поскольку маточное стадо мигрировало на нерестилища в впадающих реках.Однако с ухудшением условий нереста в реках нерест «летней» форели стал намного слабее, и установка рыболовных сетей в устьях рек в озеро исключила возможность естественного нереста. Таким образом, в настоящее время в реках водится молодняк, выращиваемый в инкубаториях.
В 1970 г. в уловах преобладала гегаркунинская форель — 120 т в год, за ней следовала «летняя» форель. В 1975 г. гегаркунинская форель составляла 56-70% уловов, «летняя» форель — около 20%, а «зимняя» форель и боджак составляли 7-18% (Смолей, 1979, 1979а).Это было связано с разным воздействием падения уровня воды на нерестилища каждого из подвидов. Основные нерестилища «зимней» форели высохли в 1963 году, и аквакультурам не удалось найти подходящий метод инкубации икры «зимней» форели. Хотя нерестилища «летней» форели в озере высохли намного раньше, т. Е. В 1948 году, чем у «зимней» форели, запасы оставались стабильными до 1960-х годов, поскольку маточное стадо мигрировало на нерестилища в впадающих реках.Однако с ухудшением условий нереста в реках нерест «летней» форели стал намного слабее, и установка рыболовных сетей в устьях рек в озеро исключила возможность естественного нереста. Таким образом, в настоящее время в реках водится молодняк, выращиваемый в инкубаториях.
Попытки остановить сокращение запасов форели на Севане и в реках включали введение неводов большого размера. В 1971-1973 гг. Лов форели в периоды ее интенсивного кормления был запрещен, но разрешен лов во время нереста, при котором ежегодно улавливается 50-70 т рыбы.Позже невод был запрещен. В 1976 г. пришлось прекратить промысловый промысел форели (Савваитова и др. , 1989).
В 1971-1973 гг. Лов форели в периоды ее интенсивного кормления был запрещен, но разрешен лов во время нереста, при котором ежегодно улавливается 50-70 т рыбы.Позже невод был запрещен. В 1976 г. пришлось прекратить промысловый промысел форели (Савваитова и др. , 1989).
Храмуля давала 500-600 тонн в год до конца 1950-х годов, но в последующее десятилетие уловы упали до 260 тонн в год. В 1961-1970 гг. Сети большего размера использовались в больших масштабах, и храмуля в то время составляла 33,5% от общего улова. Однако в 1971 году сетка была запрещена, потому что она вылавливала непропорционально высокий процент молоди, преимущественно самцов.Запрет помог восстановить первоначальную структуру запасов до уровня 1960-х годов (Смолей, и др. , 1985).
Доля сига в уловах неуклонно увеличивалась с начала эвтрофикации озера (Таблица 8) до 1980 г. К этому времени поголовье молодело, и в маточном стаде преобладали первые производители. Запас рыбы изменился с длинного цикла на короткий. Усилилось давление коммерческого промысла на сига, увеличилась доля более молодой рыбы в уловах.Преобладание производителей 2+ и 3+ привело к более низкой плодовитости и низкому качеству икры, что снизило скорость воспроизводства и снизило выживаемость молоди. Только 24% спелых самок нерестились в 1980 г., а скорость резорбции икры была высокой (Смолей, Южакова, 1979).
Усилилось давление коммерческого промысла на сига, увеличилась доля более молодой рыбы в уловах.Преобладание производителей 2+ и 3+ привело к более низкой плодовитости и низкому качеству икры, что снизило скорость воспроизводства и снизило выживаемость молоди. Только 24% спелых самок нерестились в 1980 г., а скорость резорбции икры была высокой (Смолей, Южакова, 1979).
4.3 Инкубаторий Производство племенного материала
Зарыбление форели из рыбоводных заводов началось в 1923 году, когда в озеро было выпущено 1,4 миллиона личинок форели.Основная цель заключалась в увеличении численности форели гегаркуни и «летней» форели. В двух форелевых хозяйствах разводили личинок в больших количествах для выпуска в реки и ручьи, питающие озеро. Начиная с 1926 г. ежегодно вылавливали 20 миллионов личинок, из которых 80% составляла форель гегаркуни. В период 1949-1956 годов, когда высохла большая часть нерестилищ форели в озере (таблица 9), среднегодовое количество выпущенных мальков форели составило 47,3 миллиона, в том числе 37,4 миллиона гегаркуни. В это время часть собранной для инкубации икры гегаркуни и «летней» форели помещали в искусственные нерестовые гнезда в малых реках.В период 1956-1970 годов на Севанских инкубаториях ежегодно производилось около 70 миллионов яиц. После вылупления мальков держали в резервуарах 2-3 недели, а затем выпускали в реки. Дадикян (1975) подсчитал, что 90% выпущенных мальков не выжили в первый месяц после выпуска. После 1975 года сбор икры форели резко сократился: за период 1975-1987 гг. Количество собранных яиц «летней» форели снизилось с 12 миллионов до 30 000, а гегаркуни — с 62 миллионов до 6,7 миллиона (таблица 10).Что касается «зимней» форели, то раньше собирали от 0,1 млн до 5 млн икры, но в 1970 году производство ее семян в инкубатории было остановлено. Стратегия управления, продиктованная уменьшением количества созревающей рыбы (таблица 11), заключалась в том, чтобы собрать меньше икры и для производства мальков массой 1-3 г. С 1978 г. вся зрелая рыба, мигрирующая в реки, использовалась исключительно для зачистки.
В это время часть собранной для инкубации икры гегаркуни и «летней» форели помещали в искусственные нерестовые гнезда в малых реках.В период 1956-1970 годов на Севанских инкубаториях ежегодно производилось около 70 миллионов яиц. После вылупления мальков держали в резервуарах 2-3 недели, а затем выпускали в реки. Дадикян (1975) подсчитал, что 90% выпущенных мальков не выжили в первый месяц после выпуска. После 1975 года сбор икры форели резко сократился: за период 1975-1987 гг. Количество собранных яиц «летней» форели снизилось с 12 миллионов до 30 000, а гегаркуни — с 62 миллионов до 6,7 миллиона (таблица 10).Что касается «зимней» форели, то раньше собирали от 0,1 млн до 5 млн икры, но в 1970 году производство ее семян в инкубатории было остановлено. Стратегия управления, продиктованная уменьшением количества созревающей рыбы (таблица 11), заключалась в том, чтобы собрать меньше икры и для производства мальков массой 1-3 г. С 1978 г. вся зрелая рыба, мигрирующая в реки, использовалась исключительно для зачистки. Эту рыбу вылавливают в устьях рек, затем очищают от икры и молок и содержат в сетчатых садках. на 10-14 дней теряют миграционный инстинкт, после чего выпускаются в озеро.Летальность в садках 2-5%. Из-за давления браконьерства невозможно оценить, имели ли выпущенные потраченные самки шанс произвести еще одну партию яиц.
Эту рыбу вылавливают в устьях рек, затем очищают от икры и молок и содержат в сетчатых садках. на 10-14 дней теряют миграционный инстинкт, после чего выпускаются в озеро.Летальность в садках 2-5%. Из-за давления браконьерства невозможно оценить, имели ли выпущенные потраченные самки шанс произвести еще одну партию яиц.
яиц храмули инкубировали в смоделированных естественных условиях еще в 1952 г. (Маркосян, и др., , 1955). Введение сосудов Вейсса значительно улучшило выживаемость инкубированных яиц, из которых в среднем вылупилось 76%, что на 37% больше, чем при использовании более ранней практики (Рубенян, 1985).
4.4 Озеро Севан как зарыбельный материал для других водоемов
Несмотря на критическое сокращение запасов форели на Севане, программа управления рыбным хозяйством по-прежнему использует озеро в качестве источника рыбы для зарыбления других водоемов. Севанская форель была завезена в Украину, Кыргызстан и Карелию. Ежегодно в прибрежных водах собирают более 10 миллионов икры сигов, которые переправляют в озера Урала, Средней Азии, Сибири и Грузии. Храмуля перенесена в водоемы Грузии, а Москва и Санкт-Петербург.Петербургские регионы.
Храмуля перенесена в водоемы Грузии, а Москва и Санкт-Петербург.Петербургские регионы.
Оплодотворенная икра форели гегаркуни была завезена в озеро Иссык-Куль (Кыргызстан) в 1930 и 1936 годах. До 1960-х годов запасы этой рыбы в озере Иссык-Куль оставались низкими из-за отсутствия подходящих нерестилищ. После того, как на реках Тон и Каракол были созданы рыбопитомники, они предоставили достаточное количество форели гегаркуни для зарыбления. В 1970-х и 1980-х годах промышленный промысел севанской форели давал 40-50 тонн в год.
В новой среде гегаркуни приобрела несколько специфических характеристик.В озере Иссык-Куль он вырастает больше, чем в озере Севан, и достигает длины 89 см и веса 17 кг по сравнению с 60 см и 4 кг на Севане. Скорость его роста в 4-6 раз выше, чем на Севане, а плодовитость в 5-6 раз выше, а количество яиц колеблется от 3300 до 17,300. На Иссык-Куле гегаркуни питаются в основном рыбой (81,8%), тогда как на Севане только самая крупная рыба была найдена, питаясь храмулей (Дадикян, 1955). Изменились и окрас, и форма тела взрослых особей. Пролетная форма гегаркуни на Иссык-Куле дала речную форму меньшего размера (Лужин, 1956).Следовательно, с момента первого появления в озере
Иссык-Куль Севанский гегаркуни претерпел значительные изменения в своей биологии. Успешную интродукцию севанской форели в озеро Иссык-Куль следует рассматривать как важный шаг к предотвращению исчезновения этого вида. В настоящее время в озере Севан он считается находящимся под большой угрозой исчезновения из-за измененной окружающей среды, которая препятствует успешному нересту этого вида. Были предприняты попытки пересадить севанскую форель в избранные небольшие водоемы в высоких горах Армении.В 1960-х годах гегаркуни выпускали в озера Кари (высота 3185 м, 12,5 га), Акна (3030 м, 50 га) и Сев (2666 м, 192 га). Однако эти озера не были подходящими нерестилищами для гегаркуни, и необходимо было регулярно зарыблять озера мальками массой 1 г. Были построены искусственные нерестилища, но результаты неизвестны.
Изменились и окрас, и форма тела взрослых особей. Пролетная форма гегаркуни на Иссык-Куле дала речную форму меньшего размера (Лужин, 1956).Следовательно, с момента первого появления в озере
Иссык-Куль Севанский гегаркуни претерпел значительные изменения в своей биологии. Успешную интродукцию севанской форели в озеро Иссык-Куль следует рассматривать как важный шаг к предотвращению исчезновения этого вида. В настоящее время в озере Севан он считается находящимся под большой угрозой исчезновения из-за измененной окружающей среды, которая препятствует успешному нересту этого вида. Были предприняты попытки пересадить севанскую форель в избранные небольшие водоемы в высоких горах Армении.В 1960-х годах гегаркуни выпускали в озера Кари (высота 3185 м, 12,5 га), Акна (3030 м, 50 га) и Сев (2666 м, 192 га). Однако эти озера не были подходящими нерестилищами для гегаркуни, и необходимо было регулярно зарыблять озера мальками массой 1 г. Были построены искусственные нерестилища, но результаты неизвестны.
Интродукция сиговых рыб из озера Севан на Иссык-Куль началась в 1965 году с передачи яиц в Тонный инкубаторий. Полученные мальки сига 3-4 суток выпускали в озеро.Всего за период 1966-1973 гг. Было выпущено 86,8 млн мальков, из них 2 млн мальков было выпущено первоначально (1966 г.), пик пришелся на 1969 г. — 20,5 млн, 17,5 млн в 1971 г., 16 млн в 1972 г. и 13,1 млн в 1973 г. (Никитин, 1976). Первые 400 кг сига были пойманы в Тюпской губе в 1974 году.
Севанский сиг считается сегодня гибридом ладожского сига и чудского сига (Шапошникова, 1971). Никитин (1976) описал морфологические различия между исходной рыбой и потомством, которое произошло от них на Иссык-Куле.Иссык-кульский сиг также имеет разные привычки питания, поскольку питается в основном бентосом (моллюски, мизиды) и гораздо меньше зоопланктоном. Скорость роста увеличилась, особенно в течение первого года жизни. В возрасте 4+ иссык-кульский сиг весит около 1,5 кг по сравнению с 1 кг севанского сига (Никитин, 1976). Самцы сига на Иссык-Куле раньше достигают половой зрелости, часто уже в возрасте 1+, а самки становятся половозрелыми в возрасте 2+. Плодовитость высокая — 27 500-50 152 яиц (Никитин, 1976).
Самцы сига на Иссык-Куле раньше достигают половой зрелости, часто уже в возрасте 1+, а самки становятся половозрелыми в возрасте 2+. Плодовитость высокая — 27 500-50 152 яиц (Никитин, 1976).
5. ДРУГИЕ ХОЛОДНЫЕ ОЗЕРА КАВКАЗА
Помимо озера Севан, ряд других озер на Кавказе имеют рыболовный потенциал. Озеро Карагель (высота 3195 м; 16,8 га), также в Армения , хотя и не имеет значения для рыболовства, имеет зарыбленную севанскую форель ( Salmo ischchan ). В озере Арпагель (2021 м; 2200 га) ведется промысел карпа, и его промысловый потенциал оценивается в 20-40 тонн в год -1 (Касымов, 1972).
В Грузия , Касымов (1972) и Чхаидзе (1981) упоминают 10 озер площадью от 30 га до 3688 га, имеющих рыбохозяйственное значение. Озеро Ханчали (высота 1930 м, площадь поверхности 1375 га) настолько мелкое, что промерзает почти до дна. Это озеро, однако, имеет потенциальную промысловую продуктивность в размере 275 тонн -1 в год, если часть будет преобразована в пруды или если будет регулярно зарыбляться. В озере Табискури (1990 м; 1440 га) водится форель и два вида сиговых. Его потенциальное производство рыбы оценивается в 100 тонн -1 в год.В озере Сагамо (1994 м; 496 га) водятся форель, карп, храмуля ( Varicorhinus capoeta ), голавль ( Leuciscus cephalus ), , сиговых и несколько других видов рыб. Потенциал производства рыбы в озере оценивается в 40 тонн -1 в год. Храмуля — коренная рыба в Грузии, она легко адаптируется к различным условиям окружающей среды и к разным типам пищи, а также имеет приятную на вкус мякоть. В Грузии он уже успешно прошел испытания в условиях аквакультуры (Верулашвили, 1981).Озеро Базалети (879 м; 124 га), где в настоящее время рыболовство отсутствует, имеет производственный потенциал 3,6 т в год -1 . Озеро Мадатапа (2108 м; 885 га) считается местом выращивания параванской расы карпа. В озере Параван (2060 м; 3680 га) водятся карп, крамуля, голавль и сига, и его рыбопродуктивный потенциал оценивается в 352 т -1 в год.
В озере Табискури (1990 м; 1440 га) водится форель и два вида сиговых. Его потенциальное производство рыбы оценивается в 100 тонн -1 в год.В озере Сагамо (1994 м; 496 га) водятся форель, карп, храмуля ( Varicorhinus capoeta ), голавль ( Leuciscus cephalus ), , сиговых и несколько других видов рыб. Потенциал производства рыбы в озере оценивается в 40 тонн -1 в год. Храмуля — коренная рыба в Грузии, она легко адаптируется к различным условиям окружающей среды и к разным типам пищи, а также имеет приятную на вкус мякоть. В Грузии он уже успешно прошел испытания в условиях аквакультуры (Верулашвили, 1981).Озеро Базалети (879 м; 124 га), где в настоящее время рыболовство отсутствует, имеет производственный потенциал 3,6 т в год -1 . Озеро Мадатапа (2108 м; 885 га) считается местом выращивания параванской расы карпа. В озере Параван (2060 м; 3680 га) водятся карп, крамуля, голавль и сига, и его рыбопродуктивный потенциал оценивается в 352 т -1 в год. Озеро Барети (1587 м; 155 га) еще не рассматривалось для рыбного промысла.
Озеро Барети (1587 м; 155 га) еще не рассматривалось для рыбного промысла.
Для эффективного управления озерами и водохранилищами в Грузии Чхаидзе (1981) подсчитал, что необходимо 38 миллионов мальков / сеголетков, в том числе 4 миллиона карпа, 10 миллионов рыб, питающихся фитопланктоном или водными макрофитами (включая 9 миллионов толстолобиков), 2 миллиона форели и 15 миллионов храмуля.В начале 80-х годов прошлого века грузинские рыбопитомники производили всего 3,5 миллиона карпа, 2 миллиона храмулей, 4-5 миллионов сиговых рыб и ни одного толстолобика.
В Абхазия Озеро Бебесини (159 га) потенциал производства рыбы оценивается в 4,7 т в год –1 (Касымов, 1972). Красноперка ( Scardinius erythrophthalmus ), окунь, линь, лещ, щука, вель, Alburnus charusini и несколько других видов рыб обитают в озере. Озеро Рица (высота 925 м) является источником форели.
За Азербайджан Касымов (1972) назвал 10 озер, имеющих рыбохозяйственное значение. Среди них на озере Гейгель (1571 м; 79 га) водится форель, которая используется для любительского рыболовства.
Среди них на озере Гейгель (1571 м; 79 га) водится форель, которая используется для любительского рыболовства.
Подводя итог: в странах Кавказа ряд озер с холодной водой, все они значительно меньше озера Севан, по-видимому, пригодны для выращивания рыбы. Основными кандидатами являются следующие виды рыб: коренная храмуля, которой в большинстве таких озер должно хватать своей любимой пищи, т. Е.е. перифитон, детрит и водоросли. Коргониды и толстолобик дополнят рыбное сообщество в качестве питателей планктона. Местная раса карпа с высокогорного озера Параван кажется подходящим дополнением к вышеперечисленным видам.
6. ОБСУЖДЕНИЕ
Озеро Севан — одно из постоянно растущего числа озер в Азии, которые подверглись серьезному стрессу в результате деятельности человека. Озеро Севан площадью 1244 км 2 является основным источником гидроэлектроэнергии.Между 1940 и 1978 годами уровень воды в ней сильно колебался в результате выработки гидроэлектроэнергии и забора воды для орошения. Опасность продолжения слива воды была признана в 1979 году, и дальнейший слив был остановлен. Однако после войны между Арменией и Азербайджаном в начале 1990-х годов, которая привела к нефтяной блокаде, осушение озера Севан для выработки электроэнергии было возобновлено. Неустойчивые гидрологические условия в сочетании с процессом эвтрофикации в результате комбинированного воздействия более мелкого озера и увеличения поступления питательных веществ из населенных пунктов вокруг озера и промышленных предприятий сильно повлияли на первоначальные рыбные запасы.Севанская форель, самый ценный вид на Севане, оказалась под серьезной угрозой исчезновения, и ее доля в выловах рыбы упала до 1% сегодня по сравнению с 60-65% до того, как вода в озере стала управляемой.
Опасность продолжения слива воды была признана в 1979 году, и дальнейший слив был остановлен. Однако после войны между Арменией и Азербайджаном в начале 1990-х годов, которая привела к нефтяной блокаде, осушение озера Севан для выработки электроэнергии было возобновлено. Неустойчивые гидрологические условия в сочетании с процессом эвтрофикации в результате комбинированного воздействия более мелкого озера и увеличения поступления питательных веществ из населенных пунктов вокруг озера и промышленных предприятий сильно повлияли на первоначальные рыбные запасы.Севанская форель, самый ценный вид на Севане, оказалась под серьезной угрозой исчезновения, и ее доля в выловах рыбы упала до 1% сегодня по сравнению с 60-65% до того, как вода в озере стала управляемой.
Был предпринят ряд мер, чтобы остановить дальнейшее ухудшение состояния озера. Больше воды было доставлено в озеро из других водосборов через несколько туннелей, что привело к повышению уровня воды в озере на 1,5 м. Запрет на ловлю севанской форели был введен в 1976 году, а национальный парк на озере Севан был создан в 1978 году. Тем не менее, ущерб продолжался, в том числе из-за продолжающегося эвтрофикации озера. После обнаружения в 1970-1973 годах придонной аноксии с присутствием сероводорода и метана в озере в 1983 году произошла массовая гибель рыбы. В 1976 году промысел форели был запрещен для защиты оставшихся запасов, но браконьерство продолжалось. запрет плохо соблюдался в период серьезных экономических ограничений (Meybeck et al. , 1997). Интродукция сиговых рыб дала начало новому промыслу, начавшемуся в 1970-х годах и завершившемуся в 1982 году с добычей 16 000 т в год.После этого произошел резкий спад.
Тем не менее, ущерб продолжался, в том числе из-за продолжающегося эвтрофикации озера. После обнаружения в 1970-1973 годах придонной аноксии с присутствием сероводорода и метана в озере в 1983 году произошла массовая гибель рыбы. В 1976 году промысел форели был запрещен для защиты оставшихся запасов, но браконьерство продолжалось. запрет плохо соблюдался в период серьезных экономических ограничений (Meybeck et al. , 1997). Интродукция сиговых рыб дала начало новому промыслу, начавшемуся в 1970-х годах и завершившемуся в 1982 году с добычей 16 000 т в год.После этого произошел резкий спад.
Падение уровня озера сократило пояс макрофитов в 30 раз, при этом резко изменилась литораль, где практически исчез Gammarus , важный источник пищи для севанской форели. Эвтрофированное озеро теперь богато зелеными и сине-зелеными водорослями, которые время от времени цветут. Прозрачность воды снизилась с 12,5 м до 4,5 м (Meybeck et al. , 1997). Хотя сейчас гидроэлектростанция включается только в чрезвычайных ситуациях, например, вызванных нефтяным эмбарго, наложенным на Армению соседним Азербайджаном в 1993 году, это вызывает дальнейшее падение уровня воды в озере Севан. Альтернативой был перезапуск устаревшей атомной электростанции для уменьшения нехватки электроэнергии в стране (Herald Tribune, 13 августа 1993 г.), что предотвратило бы дальнейшее ухудшение состояния озера и его рыбных запасов.
Альтернативой был перезапуск устаревшей атомной электростанции для уменьшения нехватки электроэнергии в стране (Herald Tribune, 13 августа 1993 г.), что предотвратило бы дальнейшее ухудшение состояния озера и его рыбных запасов.
Meybeck et al. (1997) обобщил причины почти полного исчезновения севанской форели:
- Утрата первоначальных прибрежных нерестилищ по берегам озера и в устьях впадающих рек,
- изменения качества субстрата на новой береговой литорали,
- сокращение погруженных водных макрофитов,
- изменений в кормовых организмах рыб, особенно потеря Gammarus (ракообразное, обитающее в чистой воде),
- летнее повышение температуры поверхностных вод при нересте «летней» севанской форели,
- придонная водная гипоксия,
- перелов и браконьерство,
- Низкая эффективность искусственного воспроизводства рыб,
- конкуренция между севанской форелью и завезенными экзотическими видами рыб.

Восстановление озера Севан предполагает повышение уровня озера на 6-8 м и снижение поступления биогенных веществ, что в совокупности должно снизить текущий уровень эвтрофикации и улучшить условия растворенного кислорода (Ованисян, 1994). Повышение уровня воды позволит восстановить некоторые утраченные нерестилища севанской форели, храмули и усачей. Дальнейшие нерестилища следует создавать из гравия как в озере, так и в впадающих реках. Севанская форель, перенесенная в прошлом в озеро Иссык-Куль, должна быть повторно интродуцирована, чтобы обогатить генофонд запасов, оставшихся в озере Севан.Однако считается, что озеро, вероятно, никогда не вернется в первозданное состояние.
Нынешняя сложная экономическая ситуация в Армении привела к упадку исследований и мониторинга. Meybeck et al. (1997) пришли к выводу, что возобновление регулярной научной работы и мониторинга будет зависеть от восстановления экономики страны и международной помощи.
7.
 ССЫЛКИ
ССЫЛКИАрнольди, Л.В. 1929. Донная продуктивность в озере Севан.Труды Севанской гидробиол. Станции 1 (1): 1-96). (На русском).
Баграмян, Г.А. 1984. Гидробиологический режим и технические меры для решения проблем озера Севан. В кн .: Лимнология Горных Водоемов: 17-18. Рука. ССР, Ереван. (На русском).
Чхаидзе Р.И. Фитопланктон озера Сагамо. В кн .: Рыболовные исследования внутренних водоемов Грузии: 7-13. Сборник статей. ВНИРО, Москва. (На русском).
Дадикян, М.Г. 1955. Питание севанской форели. Труды Севанской гидробиол. Станции 14: 5-76. (На русском).
Дадикян, М.Г. 1975. Биология молоди севанской форели. Биол. Ж. Армении 28 (12): 12-20. (На русском).
Дадикян, М.Г. 1986. Рыбы Армении. Рука. ССР, Ереван. 245 стр. (На русском).
Фортунатов, М.А. 1927. Однородные виды форели из озера Севан Salmo ischchan Kessler. Труды Севанской гидробиол. Станции 1: 1-131. (На русском).
Фортунатов М. А., К.Р. Фортунатова, Е.В. Куликова. 1932. Оценка природных ресурсов озера Севан. Труды Севанской гидробиол. Станции 3: 1-182. (На русском).
А., К.Р. Фортунатова, Е.В. Куликова. 1932. Оценка природных ресурсов озера Севан. Труды Севанской гидробиол. Станции 3: 1-182. (На русском).
Фридман, Г. 1950. Донная фауна озера Севан. Труды Севанской гидробиол. Станции 10: 7-92. (На русском).
Габриелян Б.К. и М. Шатуновский. 1984. Многолетние изменения плодовитости севанской храмули и факторы, ее определяющие. В кн .: Лимнология Горных Водоемов: 50-52.Рука. ССР, Ереван. (На русском).
Габриелян Б.К. и Л. Тряпицына. 1984. Многолетние изменения плодовитости севанской храмули и факторы, лежащие в основе этого. В кн .: Лимнология Горных Водоемов: 48-50. Рука. ССР, Ереван. (На русском).
Габриелян Б.К. et al. 1990. Численность и биомасса севанской храмули ( Varicorhinus capoeta sevangi Fill.) В меняющемся водоеме. Экология 2: 46-53. (На русском).
Гамбарян, М.Г. 1979. Температурный режим озера Севан. Труды Севанской гидробиол. Станции 17: 123-129. (На русском).
Гезалян, М. Г. 1979. Температурный режим озера Севан. Труды Севанской гидробиол. Станции 17: 5-23. (На русском).
Г. 1979. Температурный режим озера Севан. Труды Севанской гидробиол. Станции 17: 5-23. (На русском).
Гезалян, М.Г. и Л. Хорлашко. 1979. Кислородный режим озера Севан. Труды Севанской гидробиол. Станции 17: 24-37. (На русском).
Оганесян Р.Х. (Оганесян Р.О.). 1994. Озеро Севан вчера, сегодня.Армения Нац. Акад. Наук, Ереван. 478 стр. (На русском).
Касымов А.Г. Пресноводная фауна Кавказа. Вяз, Баку. 285 стр. (На русском).
Казарян А.Г., 1979. Фитопланктон озера Севан. Труды Севанской гидробиол. Станции 17: 75-87. (На русском).
Легович Н.А. 1979. «Цветение воды» в озере Севан в 1964-1972 гг. Труды Севанской гидробиол. Станции 17: 51-74. (На русском).
Лещинская, А.С. 1950 г.Репродуктивная биология форели озера Севан. Труды Севанской гидробиол. Станции 10: 93-175. (На русском).
Лужин, Б. 1956. Иссык-кульская форель гегаркуни. АН Киргиз. ССР, Фрунзе. 133 стр. (На русском).
Лятти, С.Я. 1932. Гидрохимические особенности озера Севан. В кн .: Материалы по исследованию Озера Севан и его Бассейна. Ереван. 101 стр. (На русском).
В кн .: Материалы по исследованию Озера Севан и его Бассейна. Ереван. 101 стр. (На русском).
Маилян Р.А. 1957. Сиг озера Севан. Труды Севанской гидробиол. Станции 15: 136-196.(На русском).
Маркосян А.Г. 1959. Добыча бентоса в озере Севан. В кн .: Труды В. И. Совещания по проблемам биологии внутренних вод: 139–145. Москва, Ленинград. (На русском).
Маркосян, А.Г. и др. 1955. Дефицит и выращивание севанской храмули. Изв. Рука. ССР 8 (7): 15-28. (На русском).
Мешкова Т. 1976. Эвтрофикация озера Севан. Биол. Журнал Армении 29 (7): 14-22. (На русском).
Мина, м.V. 1986. Микроэволюция рыб. Наука, Москва. 207pp. (На русском).
Мейбек М., М. Акопян и В. Андреасян. 1997. Что случилось с озером Севан? SIL News 23: 30 июня 1997: 7-10.
Никитин, А.А. 1976. Интродукция и искусственное выращивание сига в водоемах Киргизии. Илим, Фрунзе. 121 стр. (На русском).
Никогосян, А.А. et al. 1979. Распространение зоопланктона в озере Севан. Труды Севанской гидробиол.Станции 17: 118-122. (На русском).
Распространение зоопланктона в озере Севан. Труды Севанской гидробиол.Станции 17: 118-122. (На русском).
Оганесян Р.О. и в качестве. Парпаров. 1983. Экологические аспекты проблем озера Севан. Труды Севанской гидробиол. Станции 18: 5-13. (На русском).
Оганесян Р.О., А.С. Парпаров, А.А. Симонян. 1977. Биологические основы проблем озера Севан. Биол. Ж. Армении 30 (10): 101-106. (На русском).
Островский, И.С. 1983. Продуктивность обычных видов зообентоса и их роль в экосистеме озера Севан.Москва (дипломная работа). 24стр. (На русском).
Парпаров, А.С. 1990. Некоторые характеристики сообщества автотрофов озера Севан в связи с его эвтрофикацией. Hydrobiologia 191: 15-21.
Парпарова Р.М. 1979. Гидрохимические условия озера Севан в 1976 году. Труды Севанской гидробиол. Станции 17: 38-50. (На русском).
Павлов Д.С., К.С. Савваитова, В. Маркарян. 1989. Экологический мониторинг и возможности сохранения форели Salmo ischchan (Kessler, 1877) в озере Севан. Вестник Московского университета, сер. Биол. 1: 73-78. (На русском).
Вестник Московского университета, сер. Биол. 1: 73-78. (На русском).
Павлов П.И. 1947. Нерестилища и влияние падения уровня воды на популяцию форели в озере Севан. Труды Севанской гидробиол. Станции 12: 93-140. (На русском).
Павлов П.И. 1951. Вклад в биологию севанской форели. Труды Севанской гидробиол. Станции 12: 93-140. (На русском).
Пивазян, С.А. 1972. Питание и кормовые отношения между форелью Salmo ischchan Kessler и сигом Coregonus lavaretus ladoga Поляков в озере Севан.Вопросы Ихтиол. 12 (6): 1086-1093. (На русском).
Пивазян С.А. 1979. Питание лососевых рыб озера Севан. Труды Севанской гидробиол. Станции 17: 153-161. (На русском).
Пивазян С.А. 1984. Питание и кормовые отношения между форелью и сигом в озере Севан. Труды Севанской гидробиол. Станции 19: 151-253. (На русском).
Красная книга СССР. 1984. Vol. 1. Лесная промышленность, Москва. 390pp. (На русском).
Рубенян, А.Р. 1985. Способ искусственного выращивания севанской храмули. Труды Севанской гидробиол. Станции 20: 245-253. (На русском).
1985. Способ искусственного выращивания севанской храмули. Труды Севанской гидробиол. Станции 20: 245-253. (На русском).
Савваитова К.А., Э.А. Дорофеева, В. Маркарян, А.И. Смолей. 1989. Форель озера Севан. Труды Зоол. Inst. АН СССР (Ленинград). Vol. 204. 180 стр. (На русском).
Шапошникова, Г.Х. 1971. Сравнительная морфология рода Coregonus из озера Севан. Вопросы Ихтиол. 11 (4): 575-586. (На русском).
Симонян, А.А. 1988. Зоопланктон в изменяющихся условиях водоема (на примере озера Севан). Ленинград (диссертация). 43 стр. (На русском).
Смолей, А. 1979. Биология и численность севанской форели в условиях изменения озерных условий. Труды Севанской гидробиол. Станции 17: 181-211. (На русском).
Смолей, А. 1979а. Динамика численности лососевых рыб в озере Севан в изменяющейся среде. Труды Севанской гидробиол. Станции 17: 221-227. (На русском).
Смолей, А.I. 1986. Биологические аспекты увеличения численности севанской форели. В кн .: Динамика численности промышленных. Рыб: 160-166. Наука, Москва. (На русском).
В кн .: Динамика численности промышленных. Рыб: 160-166. Наука, Москва. (На русском).
Смолей А.И., Пивазян С. Южакова. 1985. Рыбные ресурсы озера Севан во время падения уровня воды и возможности их эксплуатации. Труды Севанской гидробиол. Станции 20: 199-244. (На русском).
Смолей, А. и Г. Южакова. 1979. Обилие лососевых в озере Севан в 1971-1975 гг.Труды Севанской гидробиол. Станции 17: 212-220. (На русском).
Южакова Г.Г. 1979. Обилие севанского сига в 1961-1970 гг. Труды Севанской гидробиол. Станции 17: 172-181. (На русском).
Южакова Г.Г. 1985. Рост и половое созревание севанского сига. Труды Севанской гидробиол. Станции 20: 188-198. (На русском).
Южакова Г.Г. и С.А.Пивазян. 1975. Обилие интродуцированного сига в озере Севан. Изв. Государь.Науч.-исслед. Inst. Озер. Речь. Рыб. Хоз. (ГОСНИОРХ), Ленинград, 103: 202-206. (На русском).
Верулашвили, Г.Г. 1981. Храмуля ( Varicorhinus capoeta ), полезная рыба для рыболовства. В: Рыболовные исследования внутренних водоемов Грузии: 65-69. ВНИРО, Москва. (На русском).
В: Рыболовные исследования внутренних водоемов Грузии: 65-69. ВНИРО, Москва. (На русском).
Аутоиммунный гепатит: основы практики, история вопроса, патофизиология
Эттингер Р., Бруннберг А., Гернер П., Винтермейер П., Дженке А., Вирт С.Клинические особенности и биохимические данные у детей европеоидной расы при диагностике аутоиммунного гепатита. J Аутоиммунный . 2005 24 февраля (1): 79-84. [Медлайн].
Strassburg CP, Manns MP. Лечение аутоиммунного гепатита. Semin Liver Dis . 2009 29 августа (3): 273-85. [Медлайн].
[Рекомендации] Мак К.Л., Адамс Д., Ассис Д.Н. и др. Диагностика и лечение аутоиммунного гепатита у взрослых и детей: практическое руководство и рекомендации 2019 года Американской ассоциации по изучению заболеваний печени. Гепатология . 2020 72 августа (2): 671-722. [Медлайн]. [Полный текст].
[Медлайн]. [Полный текст].
Montano-Loza AJ, Carpenter HA, Czaja AJ. Последствия отмены лечения при аутоиммунном гепатите 1 типа. Печень Инт . 2007 май. 27 (4): 507-15. [Медлайн].
Waldenstrom J. Диагностическое значение АКТГ. Acta Endocrinol (Копенг) . 1950. 5 (3): 235-42. [Медлайн].
Kunkel HG, Аренс EH, Eisenmenger WJ.Сильная гипергаммаглобулинемия у молодых женщин с заболеванием печени неустановленной этиологии. Дж. Клин Инвест . 1950. 30: 654.
Bearn AG, Kunkel HG, Slater RJ. Проблемы хронических заболеваний печени у молодых женщин. Am J Med . 1956. 21: 3-15.
Joske RA, King WE. Феномен L.E.-клеток при активном хроническом вирусном гепатите. Ланцет . 1955, 3 сентября. 269 (6888): 477-80. [Медлайн].
Ланцет . 1955, 3 сентября. 269 (6888): 477-80. [Медлайн].
Обтекатель DC, Маккей IR, Taft LI.Лупоидный гепатит. Ланцет . 1956 29 декабря. 271 (6957): 1323-6. [Медлайн].
Джонсон П.Дж., Макфарлейн И.Г. Отчет о встрече: Международная группа по аутоиммунным гепатитам. Гепатология . 1993 18 октября (4): 998-1005. [Медлайн].
Скалли Л.Дж., Тозе С., Сенгар Д.П. и др. Ранний аутоиммунный гепатит связан с делецией гена C4A. Гастроэнтерология . 1993 Май. 104 (5): 1478-84. [Медлайн].
Czaja AJ, Carpenter HA, Santrach PJ, et al.Генетическая предрасположенность иммунологических особенностей хронического активного гепатита. Гепатология . 1993 18 октября (4): 816-22. [Медлайн].
 webmd.com»> Oo YH, Hubscher SG, Adams DH. Аутоиммунный гепатит: новые парадигмы в патогенезе, диагностике и лечении. Гепатол Инт . 2010 19 мая. 4 (2): 475-93. [Медлайн]. [Полный текст].
webmd.com»> Oo YH, Hubscher SG, Adams DH. Аутоиммунный гепатит: новые парадигмы в патогенезе, диагностике и лечении. Гепатол Инт . 2010 19 мая. 4 (2): 475-93. [Медлайн]. [Полный текст].
Флореани А., Рестрепо-Хименес П., Секки М.Ф. и др. Этиопатогенез аутоиммунного гепатита. J Аутоиммунный .2018 декабрь 95: 133-43. [Медлайн].
Умемура Т., Ота М. Генетические факторы влияют на этиологию, клинические характеристики и исход аутоиммунного гепатита. Clin J Гастроэнтерол . 2015 8 (6): 360-6. [Медлайн].
Ван К., Ян Ф., Мяо К., Кравитт Е. Л., Гершвин М. Е., Ма X. Клинические фенотипы аутоиммунного гепатита: всесторонний обзор. J Аутоиммунный . 2016, январь, 66: 98-107. [Медлайн].
Czaja AJ.Диагностика и лечение аутоиммунного гепатита. Clin Liver Dis . 2015 19 февраля (1): 57-79. [Медлайн].
Clin Liver Dis . 2015 19 февраля (1): 57-79. [Медлайн].
Pathmakanthan S, Kay EW, Murray FE. Аутоиммунный хронический активный гепатит, связанный с наличием антифосфолипидных антител. Eur J Гастроэнтерол Hepatol . 1998 10 февраля (2): 155-7. [Медлайн].
Ben-Ari Z, Czaja AJ. Аутоиммунный гепатит и его вариантные синдромы. Кишечник . 2001 Октябрь, 49 (4): 589-94.[Медлайн]. [Полный текст].
Czaja AJ. Аутоантител-отрицательный аутоиммунный гепатит. Dig Dis Sci . 2012 Март 57 (3): 610-24. [Медлайн].
Czaja AJ. Аутоиммунный гепатит. Feldman M, Scharschmidt BF, Sleisenger MH, ред. Болезни желудочно-кишечного тракта и печени Слейзенгера и Фордтрана . 6-е изд. Филадельфия, Пенсильвания: WB Saunders Company; 1998. 1265-74.
1265-74.
Рамакришна Дж., Джонсон А.Р., Баннер Б.Ф.Длительное применение миноциклина при акне у здоровых подростков может вызвать тяжелый аутоиммунный гепатит. Дж Клин Гастроэнтерол . 2009 Сентябрь 43 (8): 787-90. [Медлайн].
Адар Т., Мизрахи М., Паппо О., Шейман-Элазари А., Шиболет О. Аутоиммунный гепатит, индуцированный адалимумабом. Дж Клин Гастроэнтерол . 2010, январь, 44 (1): e20-2. [Медлайн].
Fairhurst DA, Sheehan-Dare R. Аутоиммунный гепатит, связанный с инфликсимабом, у пациента с ладонно-подошвенным пустулезным псориазом. Клин Экспер Дерматол . 2009 Апрель, 34 (3): 421-2. [Медлайн].
Лю ZX, Капловиц Н. Иммунно-опосредованное лекарственное заболевание печени. Clin Liver Dis . 2002 6 августа (3): 755-74. [Медлайн].
[Медлайн].
Casswall TH, Nemeth A, Nilsson I, Wadstrom T., Nilsson HO. ДНК видов Helicobacter в тканях печени и желудка у детей и подростков с хроническим заболеванием печени. Сканд Дж Гастроэнтерол . 2010. 45 (2): 160-7. [Медлайн].
Тунио Н.А., Мансур Э., Шериф М.З., Купер Г.С., Склер С.Н., Коэн С.М. Эпидемиология аутоиммунного гепатита (АИГ) в США в период с 2014 по 2019 год: национальное популяционное исследование. Дж Клин Гастроэнтерол . 2021 ноябрь-01 декабря. 55 (10): 903-10. [Медлайн].
Херлбурт К.Дж., МакМахон Б.Дж., Дойбнер Х., Хсу-Травински Б., Уильямс Д.Л., Каудли К.В. Распространенность аутоиммунного заболевания печени у коренных жителей Аляски. Ам Дж. Гастроэнтерол .2002 сентябрь 97 (9): 2402-7. [Медлайн].
 com»> Ngu JH, Bechly K, Chapman BA, et al. Популяционное эпидемиологическое исследование аутоиммунного гепатита: болезнь пожилых женщин ?. Дж Гастроэнтерол Гепатол . 2010 25 октября (10): 1681-6. [Медлайн].
com»> Ngu JH, Bechly K, Chapman BA, et al. Популяционное эпидемиологическое исследование аутоиммунного гепатита: болезнь пожилых женщин ?. Дж Гастроэнтерол Гепатол . 2010 25 октября (10): 1681-6. [Медлайн].
Czaja AJ. Особые клинические проблемы при аутоиммунном гепатите: пожилые люди, мужчины, беременность, легкое заболевание, молниеносное начало и небелые пациенты. Semin Liver Dis . 2009 29 августа (3): 315-30.[Медлайн].
Миели-Вергани Г., Вергани Д. Аутоиммунный гепатит у детей: чем отличается от АИГ у взрослых ?. Semin Liver Dis . 2009 29 августа (3): 297-306. [Медлайн].
Хайдер А.С., Кей Дж., Томсон А. Аутоиммунный гепатит в демографически изолированном районе Австралии. Intern Med J . 2010 Апрель 40 (4): 281-5. [Медлайн].
 webmd.com»> Czaja AJ, Carpenter HA. Отличительный клинический фенотип и исход лечения аутоиммунного гепатита 1 типа у пожилых. Гепатология . 2006 Март 43 (3): 532-8. [Медлайн].
webmd.com»> Czaja AJ, Carpenter HA. Отличительный клинический фенотип и исход лечения аутоиммунного гепатита 1 типа у пожилых. Гепатология . 2006 Март 43 (3): 532-8. [Медлайн].
Montano-Loza AJ, Carpenter HA, Czaja AJ. Особенности, связанные с неэффективностью лечения аутоиммунного гепатита 1 типа, и прогностическая ценность модели терминальной стадии заболевания печени. Гепатология . 2007 Октябрь 46 (4): 1138-45. [Медлайн].
Грегорио Г.В., Портманн Б., Рид Ф. и др. Аутоиммунный гепатит в детстве: 20-летний опыт. Гепатология . 1997 Март.25 (3): 541-7. [Медлайн].
Mieli-Vergani G, Vergani D, Baumann U, et al. Диагностика и лечение аутоиммунного заболевания печени у детей: Заявление о позиции комитета по гепатологии ESPGHAN. J Педиатр Гастроэнтерол Нутр . 2018 Февраль 66 (2): 345-60. [Медлайн]. [Полный текст].
[Медлайн]. [Полный текст].
[Рекомендации] Глисон Д., Хенеган М.А., Британское общество гастроэнтерологов. Рекомендации Британского общества гастроэнтерологов по лечению аутоиммунного гепатита. Кишечник . 2011 Декабрь 60 (12): 1611-29. [Медлайн].
Buechter M, Manka P, Heinemann FM, et al. Потенциальные провоцирующие факторы острой печеночной недостаточности как первого проявления аутоиммунного гепатита — единый опыт исследования 52 взрослых пациентов. Мир Дж. Гастроэнтерол . 2018 7 апреля. 24 (13): 1410-8. [Медлайн]. [Полный текст].
Кирк А.П., Джейн С., Покок С. и др. Поздние результаты проспективного контролируемого исследования Royal Free Hospital по терапии преднизолоном при хроническом активном гепатите, отрицательном по поверхностному антигену. Кишечник . 1980, 21 января (1): 78-83. [Медлайн].
[Медлайн].
Czaja AJ, Rakela J, Ludwig J. Особенности, отражающие ранний прогноз тяжелого аутоиммунного хронического активного гепатита, принимающего кортикостероиды. Гастроэнтерология . 1988 Август 95 (2): 448-53. [Медлайн].
Roberts SK, Therneau TM, Czaja AJ. Прогноз гистологического цирроза печени при аутоиммунном гепатите 1 типа. Гастроэнтерология . 1996 Mar.110 (3): 848-57. [Медлайн].
Ferreira AR, Roquete ML, Toppa NH, de Castro LP, Fagundes ED, Penna FJ. Эффект лечения гистопатологии печени у детей и подростков с аутоиммунным гепатитом. J Педиатр Гастроэнтерол Нутр . 2008, январь, 46 (1): 65-70. [Медлайн].
Czaja AJ, Menon KV, Carpenter HA. Устойчивая ремиссия после терапии кортикостероидами аутоиммунного гепатита 1 типа: ретроспективный анализ.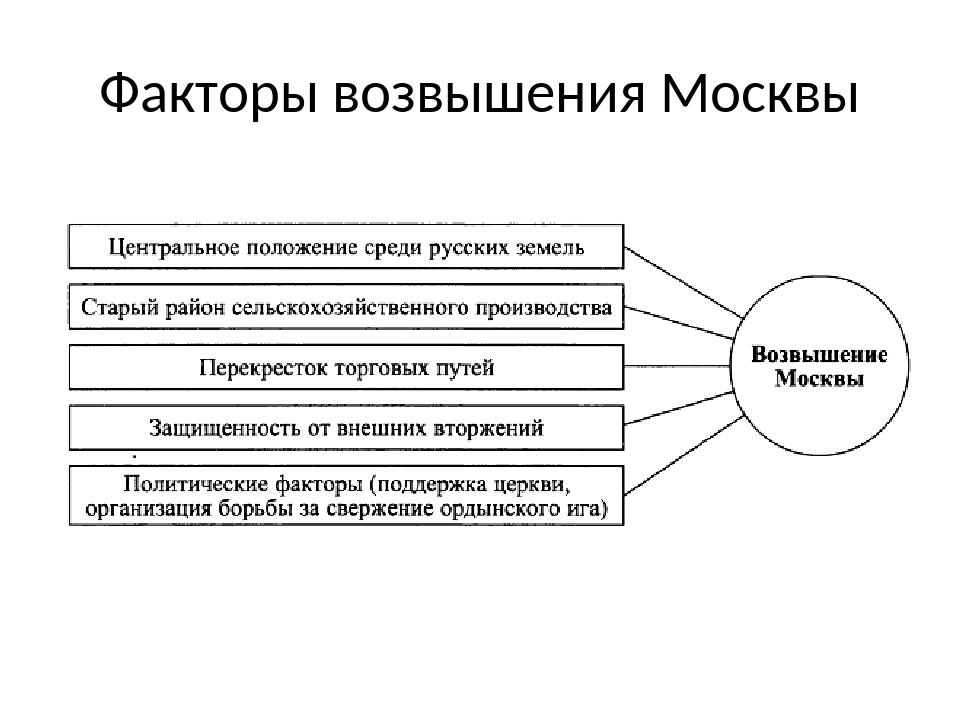 Гепатология . 2002 апр.35 (4): 890-7. [Медлайн].
Гепатология . 2002 апр.35 (4): 890-7. [Медлайн].
[Рекомендации] Manns MP, Czaja AJ, Gorham JD, et al. Диагностика и лечение аутоиммунного гепатита. Гепатология . 2010 июн. 51 (6): 2193-213. [Медлайн].
Hoeroldt B, McFarlane E, Dube A и др. Отдаленные результаты лечения пациентов с аутоиммунным гепатитом в нетрансплантационном центре. Гастроэнтерология . 2011 июн. 140 (7): 1980-9. [Медлайн]. [Полный текст].
Глисон Д.Отдаленные исходы аутоиммунного гепатита. Clin Liver Dis (Хобокен) . 14 июля 2019 г. (1): 24-8. [Медлайн]. [Полный текст].
Рахим М.Н., Микель Р., Хенеган Массачусетс. Обращение к пациенту с острым тяжелым аутоиммунным гепатитом. Представитель JHEP . 2020 Декабрь 2 (6): 100149. [Медлайн]. [Полный текст].
[Полный текст].
Шарма С., Агарвал С., Каушал К. и др. Наличие и тип декомпенсации влияет на исходы аутоиммунного гепатита при лечении кортикостероидами. JGH Открыть . 2021 5 января (1): 81-90. [Медлайн]. [Полный текст].
Caprai S, Vajro P, Ventura A, et al. Аутоиммунное заболевание печени, связанное с глютеновой болезнью в детстве: многоцентровое исследование. Клин Гастроэнтерол Гепатол . 2008 июл.6 (7): 803-6. [Медлайн].
Миели-Вергани Г., Вергани Д. Будесонид при ювенильном аутоиммунном гепатите? Еще нет. Дж. Педиатр . 2013 ноябрь 163 (5): 1246-8. [Медлайн]. [Полный текст].
Грин MT, Уитингтон PF. Исходы при аутоиммунном гепатите у детей. Curr Gastroenterol Rep . 2009 июн.11 (3): 248-51. [Медлайн].
 webmd.com»> Миели-Вергани Г., Вергани Д. Аутоиммунный гепатит в детском возрасте. Clin Liver Dis (Хобокен) . 2014 г., 3 (1): 6-8. [Медлайн]. [Полный текст].
webmd.com»> Миели-Вергани Г., Вергани Д. Аутоиммунный гепатит в детском возрасте. Clin Liver Dis (Хобокен) . 2014 г., 3 (1): 6-8. [Медлайн]. [Полный текст].
Миели-Вергани Г., Вергани Д. Аутоиммунное заболевание печени у детей. Мир Дж. Гастроэнтерол . 2008 7 июн.14 (21): 3360-7. [Медлайн]. [Полный текст].
Марсо Г., Лапьер П., Беланд К., Судейнс Х., Альварес Ф. Аутоантитела к LKM1 при хронической инфекции гепатита С: случай молекулярной мимикрии ?. Гепатология . 2005 Сентябрь 42 (3): 675-82. [Медлайн]. [Полный текст].
Frenzel C, Herkel J, Luth S, Galle PR, Schramm C, Lohse AW. Оценка F-actin ELISA для диагностики аутоиммунного гепатита. Ам Дж. Гастроэнтерол . 2006 декабрь 101 (12): 2731-6.[Медлайн].
Czaja AJ, Carpenter HA. Чувствительность, специфичность и предсказуемость интерпретации биопсии при хроническом гепатите. Гастроэнтерология . 1993 декабрь 105 (6): 1824-32. [Медлайн].
Чувствительность, специфичность и предсказуемость интерпретации биопсии при хроническом гепатите. Гастроэнтерология . 1993 декабрь 105 (6): 1824-32. [Медлайн].
Hennes EM, Zeniya M, Czaja AJ, et al, для Международной группы по аутоиммунным гепатитам. Упрощенные критерии диагностики аутоиммунного гепатита. Гепатология . 2008 июл. 48 (1): 169-76. [Медлайн]. [Полный текст].
Альварес Ф., Берг П.А., Бьянки Ф. Б. и др.Отчет Международной группы по аутоиммунному гепатиту: обзор критериев диагностики аутоиммунного гепатита. Дж. Hepatol . 1999, 31 ноября (5): 929-38. [Медлайн].
Wiegard C, Schramm C, Lohse AW. Системы оценки для диагностики аутоиммунного гепатита: прошлое, настоящее и будущее. Semin Liver Dis . 2009 29 августа (3): 254-61. [Медлайн].
 webmd.com»> Muratori P, Lalanne C, Barbato E, et al. Особенности и прогрессирование бессимптомного аутоиммунного гепатита в Италии. Клин Гастроэнтерол Гепатол . 2016 14 января (1): 139-46. [Медлайн].
webmd.com»> Muratori P, Lalanne C, Barbato E, et al. Особенности и прогрессирование бессимптомного аутоиммунного гепатита в Италии. Клин Гастроэнтерол Гепатол . 2016 14 января (1): 139-46. [Медлайн].
Manns MP, Woynarowski M, Kreisel W, et al. В контролируемом исследовании пациентов с аутоиммунным гепатитом будесонид вызывает ремиссию более эффективно, чем преднизон. Гастроэнтерология . 2010 Октябрь 139 (4): 1198-206. [Медлайн].
Woynarowski M, Nemeth A, Baruch Y, et al, для Европейской исследовательской группы по аутоиммунному гепатиту и будесониду. Будесонид в сравнении с преднизоном с азатиоприном для лечения аутоиммунного гепатита у детей и подростков. Дж. Педиатр . 2013 ноябрь 163 (5): 1347-53.e1. [Медлайн].
Schoon EJ, Bollani S, Mills PR и др., Для исследовательской группы Matrix. Минеральная плотность костной ткани в зависимости от эффективности и побочных эффектов будесонида и преднизолона при болезни Крона. Клин Гастроэнтерол Гепатол . 2005 Февраль 3 (2): 113-21. [Медлайн]. [Полный текст].
Минеральная плотность костной ткани в зависимости от эффективности и побочных эффектов будесонида и преднизолона при болезни Крона. Клин Гастроэнтерол Гепатол . 2005 Февраль 3 (2): 113-21. [Медлайн]. [Полный текст].
Zachou K, Gatselis N, Papadamou G, Rigopoulou EI, Dalekos GN. Микофенолат для лечения аутоиммунного гепатита: проспективная оценка его эффективности и безопасности для индукции и поддержания ремиссии в большой группе пациентов, не получавших лечения. Дж. Hepatol . 2011 Сентябрь 55 (3): 636-46. [Медлайн].
Альварес Ф., Чокка М., Канеро-Веласко С. и др. Кратковременное применение циклоспорина вызывает ремиссию аутоиммунного гепатита у детей. Дж. Hepatol . 1999 Февраль 30 (2): 222-7. [Медлайн].
Sciveres M, Caprai S, Palla G, Ughi C, Maggiore G. Эффективность и безопасность циклоспорина в качестве терапии аутоиммунных заболеваний печени у детей и подростков. Алимент Фармакол Тер .2004 15 января. 19 (2): 209-17. [Медлайн].
Алимент Фармакол Тер .2004 15 января. 19 (2): 209-17. [Медлайн].
Ван Тиль Д.Х., Райт Х., Кэрролл П. и др. Такролимус: потенциально новое средство для лечения аутоиммунного хронического активного гепатита: результаты открытого предварительного исследования. Ам Дж. Гастроэнтерол . 1995 Май. 90 (5): 771-6. [Медлайн].
Chazouilleres O, Wendum D, Serfaty L, et al. Долгосрочные результаты и ответ на терапию синдрома перекрытия первичного билиарного цирроза и аутоиммунного гепатита. Дж. Hepatol .2006 Февраль 44 (2): 400-6. [Медлайн].
Floreani A, Rizzotto ER, Ferrara F, et al. Клиническое течение и исходы синдрома наложения аутоиммунного гепатита и первичного склерозирующего холангита. Ам Дж. Гастроэнтерол . 2005 июл.100 (7): 1516-22. [Медлайн].
 webmd.com»> Czaja AJ, Carpenter HA. Гистологические особенности, связанные с рецидивом после отмены кортикостероидов при аутоиммунном гепатите 1 типа. Печень Инт . 2003 апреля 23 (2): 116-23.[Медлайн].
webmd.com»> Czaja AJ, Carpenter HA. Гистологические особенности, связанные с рецидивом после отмены кортикостероидов при аутоиммунном гепатите 1 типа. Печень Инт . 2003 апреля 23 (2): 116-23.[Медлайн].
Guirguis J, Alonso Y, Lopez R, Carey W. Хорошо контролируемая отмена лечения аутоиммунного гепатита может быть безопасно завершена без руководства по биопсии печени. Гастроэнтерол Реп (Oxf) . 2018 6 ноября (4): 284-90. [Медлайн]. [Полный текст].
Джонсон П.Дж., Макфарлейн И.Г., Уильямс Р. Азатиоприн для длительного поддержания ремиссии аутоиммунного гепатита. N Engl J Med . 1995, 12 октября. 333 (15): 958-63. [Медлайн].
Fernandes NF, Redeker AG, Vierling JM и др. Терапия циклоспорином у пациентов с аутоиммунным гепатитом, устойчивым к стероидам. Ам Дж. Гастроэнтерол . 1999, январь, 94 (1): 241-8. [Медлайн].
1999, январь, 94 (1): 241-8. [Медлайн].
Aqel BA, Machicao V, Rosser B и др. Эффективность такролимуса при лечении стероидорезистентного аутоиммунного гепатита. Дж Клин Гастроэнтерол . 2004 Октябрь 38 (9): 805-9. [Медлайн].
Девлин С.М., Суэйн М.Г., Урбански С.Дж. и др.Микофенолят мофетил для лечения аутоиммунного гепатита у пациентов, невосприимчивых к стандартной терапии. Банка Дж Гастроэнтерол . 2004 г., май. 18 (5): 321-6. [Медлайн].
Inductivo-Yu I, Adams A, Gish RG, et al. Микофенолят мофетил у пациентов с аутоиммунным гепатитом, не отвечающих на стандартную иммуносупрессивную терапию или не переносящих ее. Клин Гастроэнтерол Гепатол . 2007 июл.5 (7): 799-802. [Медлайн].
Ричардсон Полицейский, Джеймс Полицейский, Райдер SD. Микофенолят мофетил для поддержания ремиссии аутоиммунного гепатита у пациентов, резистентных к азатиоприну или непереносимых к нему. Дж. Hepatol . 2000 Сентябрь 33 (3): 371-5. [Медлайн].
Микофенолят мофетил для поддержания ремиссии аутоиммунного гепатита у пациентов, резистентных к азатиоприну или непереносимых к нему. Дж. Hepatol . 2000 Сентябрь 33 (3): 371-5. [Медлайн].
Вольф Д.К., Бохито Л., Фаччиуто М., Лебовикс Э. Микофенолят мофетил при аутоиммунном гепатите: единый практический опыт. Dig Dis Sci . 2009 ноябрь 54 (11): 2519-22. [Медлайн].
Aw MM, Dhawan A, Samyn M, Bargiota A, Mieli-Vergani G.Микофенолят мофетил как средство экстренной помощи при аутоиммунном заболевании печени у детей: наблюдение в течение 5 лет. Дж. Hepatol . 2009 Июль 51 (1): 156-60. [Медлайн].
Than NN, Wiegard C, Weiler-Normann C, et al. Длительное наблюдение пациентов с трудно поддающимся лечению аутоиммунным гепатитом 1 типа, получающих терапию такролимусом. Сканд Дж Гастроэнтерол . 2016 Март 51 (3): 329-36. [Медлайн].
2016 Март 51 (3): 329-36. [Медлайн].
Остерман М.Т., Кунду Р., Лихтенштейн Г.Р. и др.Связь уровней нуклеотидов 6-тиогуанина и активности воспалительного заболевания кишечника: метаанализ. Гастроэнтерология . 2006 Апрель 130 (4): 1047-53. [Медлайн].
Wright HL, Bou-Abboud CF, Hassanein T, et al. Рецидив заболевания и отторжение после трансплантации печени по поводу аутоиммунного хронического активного заболевания печени. Трансплантация . 1992, январь, 53 (1): 136-9. [Медлайн]. [Полный текст].
Монтано-Лоза А.Дж., Мейсон А.Л., Ма М., Бастиампиллай Р.Дж., Бейн В.Г., Тандон П.Факторы риска рецидива аутоиммунного гепатита после трансплантации печени. Трансплантация печени . 2009 29 сентября, 15 (10): 1254-1261. [Медлайн].
 com»> Tripathi D, Neuberger J. Аутоиммунный гепатит и трансплантация печени: показания, результаты и лечение рецидивирующего заболевания. Semin Liver Dis . 2009 29 августа (3): 286-96. [Медлайн].
com»> Tripathi D, Neuberger J. Аутоиммунный гепатит и трансплантация печени: показания, результаты и лечение рецидивирующего заболевания. Semin Liver Dis . 2009 29 августа (3): 286-96. [Медлайн].
Molmenti EP, Netto GJ, Murray NG, et al. Заболеваемость и рецидивы аутоиммунного / аллоиммунного гепатита у реципиентов трансплантата печени. Трансплантация печени . 2002 июн. 8 (6): 519-26. [Медлайн]. [Полный текст].
Пост ди-джея, Дуглас Д. Д., Маллиган, округ Колумбия. Иммуносупрессия при трансплантации печени. Трансплантация печени . 2005 11 ноября (11): 1307-14. [Медлайн]. [Полный текст].
Центры по контролю и профилактике заболеваний. Рекомендации по выявлению хронической инфекции вируса гепатита С у лиц 1945-1965 годов рождения. MMWR Recomm Rep . 2012 17 августа.61: 1-32. [Медлайн]. [Полный текст].
[Медлайн]. [Полный текст].
Longhi MS, Ma Y, Mieli-Vergani G, Vergani D. Этиопатогенез аутоиммунного гепатита. J Аутоиммунный . 2010 Февраль 34 (1): 7-14. [Медлайн].
Vento S, Cainelli F. Есть ли роль вирусов в запуске аутоиммунного гепатита ?. Аутоиммунная Ред. . 2004 г., 3 (1): 61-9. [Медлайн].
Вен Л., Пикман М., Лобо-Йео А. и др. Направленное Т-клетками повреждение гепатоцитов при аутоиммунном хроническом активном гепатите. Ланцет . 1990 22-29 декабря. 336 (8730): 1527-30. [Медлайн].
Michitaka K, Nishiguchi S, Aoyagi Y, et al, для Японской группы по изучению этиологии цирроза печени. Этиология цирроза печени в Японии: общенациональное исследование. Дж Гастроэнтерол . 2010. 45 (1): 86-94. [Медлайн].
 webmd.com»> Секи Т., Кийосава К., Иноко Х. и др. Ассоциация аутоиммунного гепатита с HLA-Bw54 и DR4 у японских пациентов. Гепатология .1990 декабря 12 (6): 1300-4. [Медлайн].
webmd.com»> Секи Т., Кийосава К., Иноко Х. и др. Ассоциация аутоиммунного гепатита с HLA-Bw54 и DR4 у японских пациентов. Гепатология .1990 декабря 12 (6): 1300-4. [Медлайн].
Боберг КМ. Распространенность и эпидемиология аутоиммунного гепатита. Clin Liver Dis . 2002 6 августа (3): 635-47. [Медлайн].
Макфарлейн И.Г., Хенеган Массачусетс. Аутоиммунитет и женская печень. Hepatol Res . 2004 г., 28 (4): 171-6. [Медлайн].
Czaja AJ. Аутоиммунный гепатит и вирусная инфекция. Гастроэнтерол Clin North Am . 1994 23 сентября (3): 547-66.[Медлайн].
Czaja AJ, Cassani F, Cataleta M, Valentini P, Bianchi FB. Частота и значение антител к актину при аутоиммунном гепатите 1 типа. Гепатология . 1996 24 ноября (5): 1068-73. [Медлайн].
[Медлайн].
Такер С.М., Йонас М.М., Перес-Атайде АР. Гиалиновые капли в клетках Купфера: новый диагностический ключ для аутоиммунного гепатита. Am J Surg Pathol . 2015 июн. 39 (6): 772-8. [Медлайн].
Лихтенштейн ГР.Использование лабораторных исследований в качестве руководства по терапии 6-меркаптопурином / азатиоприном. Гастроэнтерология . 2004 ноябрь 127 (5): 1558-64. [Медлайн].
Czaja AJ, Lindor KD. Неудача будесонида в пилотном исследовании лечебно-зависимого аутоиммунного гепатита. Гастроэнтерология . 2000 ноябрь 119 (5): 1312-6. [Медлайн].
Zandieh I, Krygier D, Wong V и др. Использование будесонида в лечении аутоиммунного гепатита в Канаде. Банка Дж Гастроэнтерол .2008 22 апреля (4): 388-92. [Медлайн]. [Полный текст].
[Полный текст].
Faisal N, Renner EL. Рецидив аутоиммунных заболеваний печени после трансплантации печени. Мир J Hepatol . 2015 18 декабря. 7 (29): 2896-905. [Медлайн].
Reich DJ, Fiel I, Guarrera JV, et al. Трансплантация печени при аутоиммунном гепатите. Гепатология . 2000 окт. 32 (4, ч. 1): 693-700. [Медлайн].
Duclos-Vallee JC, Sebagh M, Rifai K, et al.Последующее 10-летнее исследование пациентов, перенесших трансплантацию по поводу аутоиммунного гепатита: гистологический рецидив предшествует клиническому и биохимическому рецидиву. Кишечник . 2003 июн. 52 (6): 893-7. [Медлайн].
Дуглас Д. Будесонид полезен при детском аутоиммунном гепатите. 10 июля 2013 г. Medscape from WebMD. Доступно на https://www.medscape. com/viewarticle/807623. Доступ: 22 июля 2013 г.
com/viewarticle/807623. Доступ: 22 июля 2013 г.
[Рекомендации] Далекос Г.Н., Коскинас Дж., Папатеодоридис Г.В.Греческая ассоциация по изучению клинических рекомендаций по изучению печени: Аутоиммунный гепатит. Энн Гастроэнтерол . 2019 янв-фев. 32 (1): 1-23. [Медлайн]. [Полный текст].
Sebode M, Hartl J, Vergani D, Lohse AW, для Международной группы по аутоиммунным гепатитам (IAIHG). Аутоиммунный гепатит: от текущих знаний и клинической практики к будущим исследованиям. Печень Инт . 2018 января 38 (1): 15-22. [Медлайн].
Винницкая Е.В., Абдулхаков С.Р., Абдурахманов Д.Т. и др.Важные проблемы диагностики и лечения первичного склерозирующего холангита (на основе российского консенсуса по диагностике и лечению аутоиммунного гепатита. Москва, 2018). Тер Арх . 2019 17 марта. 91 (2): 9-15. [Медлайн].
[Медлайн].
Валота М., Тиенеманн Ф., Миссельвиц Б. Ложноположительные серологические исследования острого гепатита А и аутоиммунного гепатита у пациента с острой инфекцией вируса Эпштейна-Барра. BMJ Case Rep . 2019 10 мая. 12 (5): [Medline].
Мюррей-Лайон И.М., Стерн Р.Б., Уильямс Р.Контролируемое исследование преднизона и азатиоприна при активном хроническом гепатите. Ланцет . 1973, 7 апреля, 1 (7806): 735-7. [Медлайн].
Возвышение как избирательная сила комплексов митохондриальной дыхательной цепи ящериц Phrynocephalus на Тибетском плато
Curr Zool. 2021 Apr; 67 (2): 191–199.
Юаньтин Цзинь
1 Колледж наук о жизни, Китайский университет Цзилян, Ханчжоу, 310018, Китай
Дебора Й. К. Брандт
2 Департамент интегративной биологии Калифорнийского университета в Беркли, Беркли, Калифорния, 94720-3140, США
Цзяшенг Ли
1 Колледж наук о жизни, Китайский университет Цзилян, Ханчжоу, 310018, Китай
Юбин Ву
1 Колледж наук о жизни, Китайский университет Цзилян, Ханчжоу, 310018, Китай
Haojie Tong
1 Колледж наук о жизни, Китайский университет Цзилян, Ханчжоу, 310018, Китай
Владимир Щур
3 Международная лаборатория статистической и вычислительной геномики, Национальный исследовательский университет Высшая школа экономики, Москва, Россия
Мария Серведио, редактор Handling
1 Колледж наук о жизни, Китайский университет Цзиляна, Ханчжоу, 310018, Китай
2 Департамент интегративной биологии Калифорнийского университета в Беркли, Беркли, Калифорния, 94720-3140, США
3 Международная лаборатория статистической и вычислительной геномики, Национальный исследовательский университет «Высшая школа экономики», Москва, Россия
Авторы Юаньтин Цзинь и Дебора И. К. Брандт внесли равный вклад в эту работу
К. Брандт внесли равный вклад в эту работу
Поступила в редакцию 22 апреля 2020 г .; Принята в печать 10 сентября 2020 г.
Copyright © Автор (ы) (2020). Опубликовано Oxford University Press от имени редакции Current Zoology. Это статья в открытом доступе, распространяемая в соответствии с условиями некоммерческой лицензии Creative Commons Attribution (http://creativecommons.org/licenses/by-nc/4.0/). ), который разрешает некоммерческое повторное использование, распространение и воспроизведение на любом носителе при условии надлежащего цитирования оригинальной работы. По вопросам коммерческого повторного использования обращайтесь по адресу [email protected]. Эта статья цитируется в других статьях PMC.- Дополнительные материалы
zoaa056_Supplementary_Data.
GUID: 526C0905-ABA9-4808-AABE-376548AAD513
- Заявление о доступности данных
Данные о географических точках отбора проб и всех измеренных цветах спины и субстрата хранятся в FigShare (doi: 10.
 6084 / m. 10308572). Соответствующие авторы несут ответственность за любые личные требования к материалам.
6084 / m. 10308572). Соответствующие авторы несут ответственность за любые личные требования к материалам.
Abstract
Животные, живущие на очень больших высотах, должны адаптироваться к низким температурам и низкой доступности кислорода (гипоксия), но лежащие в основе генетические механизмы, связанные с этими адаптациями, все еще неясны.Дыхательная цепь митохондрий может обеспечивать> 95% АТФ в клетках животных, и на ее эффективность влияют температура и доступность кислорода. Следовательно, комплексы дыхательной цепи (ПКР) могут быть важными молекулярными мишенями для положительного отбора, связанного с респираторной адаптацией в условиях высокогорья. Здесь мы исследовали положительный отбор в 5 ПКР и их факторы сборки путем анализа последовательностей 106 генов, полученных с помощью РНК-секвенирования всех 15 видов китайских Phrynocephalus ящериц, которые распространены от низменностей до Тибетского плато (средняя высота> 4500 м). .Наши результаты показывают, что доказательства положительного отбора генов ПКР существенно не отличаются от факторов сборки, и мы не обнаружили разницы в давлениях отбора между 5 комплексами. Мы специально искали положительный отбор в тех линиях, где произошли изменения в высоте ареала. Группа линий, эволюционирующих от низкой к большой высоте, показывает более сильные сигналы положительного отбора, чем линии, эволюционирующие от высокой к низкой высоте. Клоны, развивающиеся от низкого к высокому уровню, также имеют больше общих кодонов при положительном отборе, хотя изменения не эквивалентны на уровне аминокислот.Это исследование продвигает наше понимание генетической основы эволюции респираторного метаболизма животных в экстремально высоких средах и предоставляет гены-кандидаты для дальнейшего подтверждения с помощью функционального анализа.
Мы специально искали положительный отбор в тех линиях, где произошли изменения в высоте ареала. Группа линий, эволюционирующих от низкой к большой высоте, показывает более сильные сигналы положительного отбора, чем линии, эволюционирующие от высокой к низкой высоте. Клоны, развивающиеся от низкого к высокому уровню, также имеют больше общих кодонов при положительном отборе, хотя изменения не эквивалентны на уровне аминокислот.Это исследование продвигает наше понимание генетической основы эволюции респираторного метаболизма животных в экстремально высоких средах и предоставляет гены-кандидаты для дальнейшего подтверждения с помощью функционального анализа.
Ключевые слова: высота , положительный отбор, рептилии, плато, комплексы дыхательной цепи
Высокая высота представляет собой проблемы для физиологии животных из-за более низких температур и доступности кислорода (Hayes and S. O’Connor 1999; Rezende and Dinizfilho 2012; Cai и другие.2013). В частности, оба эти ограничения оказывают давление на дыхательный метаболизм животных (Lachance and Tishkoff, 2013; Lim et al. , 2019). Дыхательная цепь митохондрий является важным этапом дыхательного метаболизма, обеспечивая более 95% АТФ позвоночных (Acuña-Castroviejo et al. 2001). Следовательно, комплексы митохондриальной дыхательной цепи (ПКР) являются потенциальными мишенями естественного отбора у животных, живущих в чрезвычайно высоких средах.
, 2019). Дыхательная цепь митохондрий является важным этапом дыхательного метаболизма, обеспечивая более 95% АТФ позвоночных (Acuña-Castroviejo et al. 2001). Следовательно, комплексы митохондриальной дыхательной цепи (ПКР) являются потенциальными мишенями естественного отбора у животных, живущих в чрезвычайно высоких средах.
Дыхательная цепь митохондрий состоит из 5 белковых комплексов (I-V).Эти комплексы состоят из 13 белков, кодируемых митохондриальной ДНК (мтДНК), и 84 белков, кодируемых ядерной ДНК (яДНК; дополнительная таблица S1, также см. В базе данных Комитета по номенклатуре генов HUGO (HGNC): www.genenames.org). Кроме того, растет число других белков, кодируемых яДНК, которые, как известно, участвуют в сборке этих комплексов (база данных HGNC: www.genenames.org). Недавние исследования кристаллической структуры ПКР показали, какие субъединицы (или области) играют ключевые функции, такие как канал транслокации протонов или электронов (Fiedorczuk et al.2016; Letts et al. 2016; Лобо-Ярне и Угальде 2018). Большинство белков, кодируемых мтДНК (7/13), группируются в виде гидрофобной области в составе Комплекса I (дополнительная таблица S1).
2016; Лобо-Ярне и Угальде 2018). Большинство белков, кодируемых мтДНК (7/13), группируются в виде гидрофобной области в составе Комплекса I (дополнительная таблица S1).
За последние 15–20 лет было проведено большое количество исследований, посвященных естественному отбору 13 кодируемых мтДНК генов ПКР у видов, адаптированных к высоким высотам (Zink 2005; Mishmar et al. 2006; Jin et al. 2018). Эти исследования показали доказательства различного избирательного давления на митохондриальные гены среди популяций или близкородственных видов (Schmidt et al.2001; Луо и др. 2003; Hassanin et al. 2009; Yu et al. 2011; Ли и др. 2013; Камю и др. 2017; Юань и др. 2018). Однако изучению корреляций между возвышением среды обитания и генетическими доказательствами естественного отбора (например, количеством кодонов при положительном отборе) мешал тот факт, что только несколько кодонов (обычно менее 10) кодируемых мтДНК генов ПКР являются положительно выбран. Более того, существует небольшая статистическая мощность для сравнения силы положительного отбора среди 5 ПКР, потому что меньшая часть их белков кодируется в мтДНК (13/97), и они неравномерно распределены по комплексам (например,(например, 7 из 13 белков, кодируемых мтДНК, входят в состав Комплекса I, а в Комплексе II отсутствуют белки, кодируемые мтДНК).
Phrynocephalus — важная группа ящериц, широко распространенная в западном Китае. Он подразделяется на высокогорные живородящие виды на холодные и гипоксические засушливые или полузасушливые районы Цинхай-Тибетского плато (QTP) и яйцекладущие виды, как правило, в низинных песчаных дюнах, пустынях или районах пустыни Гоби за пределами QTP (Zhao 1999). Группа, вероятно, возникла в низинах, клады разошлись около 10 миллионов лет назад и заняли высокогорья, а как низменные, так и высокогорные клады далее диверсифицировались на разные виды (Jin and Brown 2013; Solovyeva et al.2018). Максимальная разница высот между видами в живородящих и яйцекладущих группах составляет около 5000 м, а диапазон высот остается большим (∼2 307–5040 м) среди живородящих видов в пределах QTP и ∼60–1300 м для яйцекладущих видов за пределами QTP (Jin и др., 2008, 2013; Джин, Браун, 2013). Эта группа близкородственных видов, эволюционирующих в разных градиентах высоты, географически изолированных друг от друга горными преградами с небольшой площадью перекрытия (), представляет собой подходящий случай для проверки положительного отбора в ходе эволюционной истории этих видов.
Карта отбора проб из 15 Phrynocephalus экз. Местонахождение отобранных образцов показано числами, соответствующими номерам, перечисленным в дополнительной таблице S2. Ареалы видов обозначены сплошным цветом, когда они не перекрываются, а области, в которых ареалы видов перекрываются, показаны как горизонтальные цветные полосы. Карта предоставлена Центром данных по ресурсам и наукам об окружающей среде Китайской академии наук (http://www.resdc.cn/) и отредактирована с помощью ArcGIS 9.3.
Мы недавно идентифицировали 4 кодона в генах дыхательной цепи, кодируемых мтДНК, при долгосрочном положительном отборе (т.е. при отборе во всей топологии дерева) и 6 кодонов при краткосрочном положительном отборе, проанализированном с помощью моделей сайтов ветвления. (т. е. при селекции по ветвям) у Phrynocephalus ящериц (Jin et al. 2018). Мы не обнаружили перекрытия среди 6 кодонов при положительном отборе по ветвям (т.е. разные кодоны находились под отбором в каждой ветви). Интересно, что 2 кодона при выборе ответвлений соответствуют аминокислотным остаткам, ответственным за транслокацию протонов. Хотя в предыдущих исследованиях были обнаружены интересные потенциальные мишени отбора в генах дыхательной цепи, они не исследовали, как селективное давление на дыхательную цепь варьируется среди различных белковых комплексов и среди клонов Phrynocephalus , живущих на разных высотах. Чтобы повысить эффективность статистических тестов, сравнивающих выбор между группами генов и средой (высота, в случае этого исследования), важно включить гены, кодируемые яДНК, поскольку большинство белков в ПКР закодированы в ядре. .
Интересно, что 2 кодона при выборе ответвлений соответствуют аминокислотным остаткам, ответственным за транслокацию протонов. Хотя в предыдущих исследованиях были обнаружены интересные потенциальные мишени отбора в генах дыхательной цепи, они не исследовали, как селективное давление на дыхательную цепь варьируется среди различных белковых комплексов и среди клонов Phrynocephalus , живущих на разных высотах. Чтобы повысить эффективность статистических тестов, сравнивающих выбор между группами генов и средой (высота, в случае этого исследования), важно включить гены, кодируемые яДНК, поскольку большинство белков в ПКР закодированы в ядре. .
На сегодняшний день многие исследования показали, что митохондриальные гены могут быть положительно отобраны в соответствии с градиентами окружающей среды (Fonseca et al. 2008; Foote et al. 2011; Zhou et al. 2014; Ngatia et al. 2019). Недавно несколько исследований с использованием 13 генов ПКР митохондриальных геномов показали, что высотный или продольный ассоциированный отбор происходил в разных группах животных (Луо и др. , 2003; Мишмар и др., 2006; Фонсека и др., 2008; Джин и др., 2018). Некоторые из этих исследований показали, что Комплекс I находится под более сильным селективным давлением, чем другие комплексы (Mishmar et al.2006; Yu et al. 2011; Jin et al. 2018). Однако эта гипотеза никогда не могла быть формально проверена только с использованием митохондриальных генов по следующим причинам: 1) в митогеноме кодируется слишком мало генов-кандидатов (только 13), и они неравномерно распределены между 5 ПКР, например, существует в составе Комплекса II отсутствует кодируемый мтДНК белок, и большинство белков, кодируемых мтДНК (7 из 13), были сгруппированы исключительно как гидрофобная область Комплекса I; 2) было обнаружено слишком мало выбранных кодонов, что статистически затрудняет сравнение давления отбора между ветвями.
, 2003; Мишмар и др., 2006; Фонсека и др., 2008; Джин и др., 2018). Некоторые из этих исследований показали, что Комплекс I находится под более сильным селективным давлением, чем другие комплексы (Mishmar et al.2006; Yu et al. 2011; Jin et al. 2018). Однако эта гипотеза никогда не могла быть формально проверена только с использованием митохондриальных генов по следующим причинам: 1) в митогеноме кодируется слишком мало генов-кандидатов (только 13), и они неравномерно распределены между 5 ПКР, например, существует в составе Комплекса II отсутствует кодируемый мтДНК белок, и большинство белков, кодируемых мтДНК (7 из 13), были сгруппированы исключительно как гидрофобная область Комплекса I; 2) было обнаружено слишком мало выбранных кодонов, что статистически затрудняет сравнение давления отбора между ветвями.
Чтобы преодолеть эти ограничения, мы получили кодирующую белок последовательность (CDS) ядерных генов, представляющих 5 митохондриальных ПКР всех китайских ящериц Phrynocephalus , а также сложные факторы сборки, которые функционально тесно связаны с самими ПКР. Используя эти данные, мы решаем следующие вопросы: 1) Подвергается ли какая-либо из РКС более сильному избирательному давлению, чем другие? 2) Показывают ли виды, эволюционирующие от низких к высоким высотам более сильные сигналы естественного отбора, воздействующие на их гены дыхательной цепи, чем виды, которые не претерпели таких резких изменений подъема в своей эволюционной истории? 3) Проявляют ли виды, эволюционирующие от низкого к высокому уровню, образцы параллельной эволюции на уровне кодонов или генов? Это исследование поможет выяснить генетические основы адаптации митохондриальных ПКР животных в ответ на изменения высоты.
Используя эти данные, мы решаем следующие вопросы: 1) Подвергается ли какая-либо из РКС более сильному избирательному давлению, чем другие? 2) Показывают ли виды, эволюционирующие от низких к высоким высотам более сильные сигналы естественного отбора, воздействующие на их гены дыхательной цепи, чем виды, которые не претерпели таких резких изменений подъема в своей эволюционной истории? 3) Проявляют ли виды, эволюционирующие от низкого к высокому уровню, образцы параллельной эволюции на уровне кодонов или генов? Это исследование поможет выяснить генетические основы адаптации митохондриальных ПКР животных в ответ на изменения высоты.
Материалы и методы
Образцы
В этом исследовании было использовано 15 экземпляров Phrynocephalus , собранных в 2016–2017 гг. С высот от 602 до 4541 м (дополнительная таблица S2). Были включены все 6 живородящих видов, распространенных в QTP, и 9 китайских яйцекладущих видов / линий, живущих за пределами QTP. Равные количества общей РНК, экстрагированной из каждой из 5 тканей (головного мозга, печени, мышц, сердца и легких) каждого образца, объединяли для получения библиотек последовательностей РНК. Эти виды обеспечили всесторонний охват глубоких эволюционных линий в этой группе. Все эксперименты проводились в соответствии с руководящими принципами Китайского совета по уходу за животными и были одобрены Комитетом по этике экспериментов на животных Китайского университета Цзиляна.
Эти виды обеспечили всесторонний охват глубоких эволюционных линий в этой группе. Все эксперименты проводились в соответствии с руководящими принципами Китайского совета по уходу за животными и были одобрены Комитетом по этике экспериментов на животных Китайского университета Цзиляна.
Секвенирование РНК
Всего 3 мкг РНК на образец использовали в качестве исходного материала для препаратов образцов РНК. Библиотеки секвенирования были созданы с использованием набора NEBNext ® UltraTM RNA Library Prep Kit для Illumina ® (Illumina, Сан-Диего, Калифорния, США) в соответствии с рекомендациями производителя, и к каждому образцу были добавлены коды индексов.Вкратце, мРНК очищали от общей РНК с использованием магнитных шариков, прикрепленных к поли-Т олигонуклеотидам. Фрагментацию проводили с использованием двухвалентных катионов при повышенной температуре в буфере для реакции синтеза первой цепи NEBNext (5X). КДНК первой цепи синтезировали с использованием случайного гексамерного праймера и обратной транскриптазы M-MuLV (РНКазы H-). Синтез второй цепи кДНК впоследствии выполняли с использованием ДНК-полимеразы I и РНКазы H. Остальные выступы были преобразованы в тупые концы посредством экзонуклеазной / полимеразной активности.После аденилирования 3′-концов фрагментов ДНК адаптер NEBNext со структурой петли шпильки лигировали для подготовки к гибридизации. Чтобы выбрать фрагменты кДНК длиной предпочтительно 250-300 п.н., фрагменты библиотеки очищали с помощью системы AMPure XP (Beckman Coulter, Беверли, Массачусетс, США). Затем использовали 3 мкл фермента USER (NEB, Ипсвич, Массачусетс, США) с кДНК выбранного размера, лигированной адаптером, при 37 ° C в течение 15 минут, а затем 5 минут при 95 ° C перед ПЦР. Затем проводили ПЦР с использованием ДНК-полимеразы Phusion High-Fidelity, универсальных праймеров для ПЦР и праймера Index (X).Наконец, продукты ПЦР были очищены (система AMPure XP) и качество библиотеки оценено на системе Agilent Bioanalyzer 2100.
Синтез второй цепи кДНК впоследствии выполняли с использованием ДНК-полимеразы I и РНКазы H. Остальные выступы были преобразованы в тупые концы посредством экзонуклеазной / полимеразной активности.После аденилирования 3′-концов фрагментов ДНК адаптер NEBNext со структурой петли шпильки лигировали для подготовки к гибридизации. Чтобы выбрать фрагменты кДНК длиной предпочтительно 250-300 п.н., фрагменты библиотеки очищали с помощью системы AMPure XP (Beckman Coulter, Беверли, Массачусетс, США). Затем использовали 3 мкл фермента USER (NEB, Ипсвич, Массачусетс, США) с кДНК выбранного размера, лигированной адаптером, при 37 ° C в течение 15 минут, а затем 5 минут при 95 ° C перед ПЦР. Затем проводили ПЦР с использованием ДНК-полимеразы Phusion High-Fidelity, универсальных праймеров для ПЦР и праймера Index (X).Наконец, продукты ПЦР были очищены (система AMPure XP) и качество библиотеки оценено на системе Agilent Bioanalyzer 2100.
Библиотеки секвенировали на платформе Illumina X Ten и были сгенерированы считывания с парных концов 150 п. н. Всего было получено около 10 ГБ данных РНК-секвенирования на образец.
н. Всего было получено около 10 ГБ данных РНК-секвенирования на образец.
Сборка транскриптома и контроль качества
Необработанные считывания в формате fastq были отфильтрованы путем удаления считываний, содержащих последовательности адаптеров, считываний, содержащих поли-N, и считываний низкого качества из необработанных данных.Транскриптомы de novo были собраны из каждого образца с использованием Trinity v2.5 (Grabherr et al. 2011) с параметрами –seqType fq –max_memory 60 G –CPU 20 –min_kmer_cov 3 –min_contig_length 300 –bfly_opts “-V 20.” Мы оценили качество сборки транскриптома с помощью собственного Perl-скрипта, модифицированного с https://github.com/vikas0633/perl/blob/master/CountFasta.pl. Общая длина транскриптома варьировала от 145 до 338 Мб, в среднем 174 Мб, а N50 — от 2217 до 2536, в среднем 2320.
гены ПКР и факторы сборки
Последовательности 84 генов ПКР и 41 сложный ген сборки были извлечены из 2 хорошо аннотированных эталонных геномов рептилий (ящерица: Anolis carolinensis ; змея: Ophiophagus hannah ).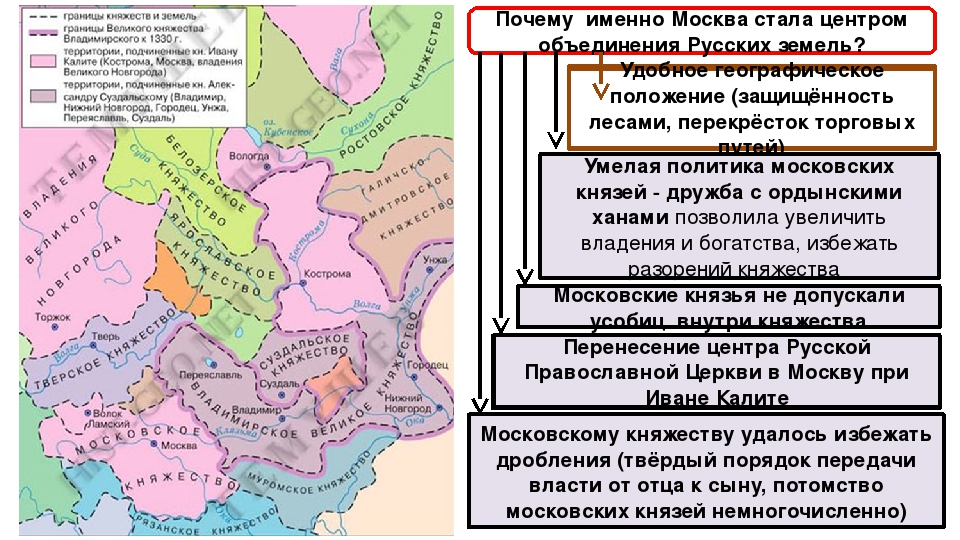 Контиги транскриптомов были выровнены с этими эталонными последовательностями с помощью blastx, и мы выбрали самый длинный транскрипт из каждого вида Phrynocephalus , который выровняли с каждым из 125 генов для использования в этом исследовании. Мы использовали только одну репрезентативную последовательность для каждого гена и для каждого вида.Поэтому мы отмечаем, что в наших данных не было гетерозиготных сайтов, что является ограничением данного исследования. Ортологические гены были согласованы с помощью программы Kalign и отредактированы вручную. Каждый транскрипт искали в базе данных неизбыточных белковых последовательностей (nr) с помощью BlastX, и все потенциальные альтернативно сплайсированные экзоны удаляли из выравнивания перед выполнением анализа отбора. Наконец, мы исключили из нашего анализа гены, которые отсутствовали у 2 или более видов.
Контиги транскриптомов были выровнены с этими эталонными последовательностями с помощью blastx, и мы выбрали самый длинный транскрипт из каждого вида Phrynocephalus , который выровняли с каждым из 125 генов для использования в этом исследовании. Мы использовали только одну репрезентативную последовательность для каждого гена и для каждого вида.Поэтому мы отмечаем, что в наших данных не было гетерозиготных сайтов, что является ограничением данного исследования. Ортологические гены были согласованы с помощью программы Kalign и отредактированы вручную. Каждый транскрипт искали в базе данных неизбыточных белковых последовательностей (nr) с помощью BlastX, и все потенциальные альтернативно сплайсированные экзоны удаляли из выравнивания перед выполнением анализа отбора. Наконец, мы исключили из нашего анализа гены, которые отсутствовали у 2 или более видов.
Построение филогенетического дерева
Наборы данных были разделены на 3 позиции кодона для филогенетического анализа. Анализ максимального правдоподобия (ML) проводился с использованием RAxML вер. V8.2.10 в операционной системе Linux Debian. Для анализа использовалась команда raxmlHPC-PTHREADS, и для каждого раздела была указана модель GTR + G (команда -m). Дерево ML было идентифицировано для исходной последовательности, а дополнительные деревья были определены для 2000 наборов данных с повторной выборкой бутстрапов, чтобы указать уровни поддержки ветвей (опция — #). Внешняя группа не была указана, поэтому для отображения дерево машинного обучения имело корень из средней точки.
Анализ максимального правдоподобия (ML) проводился с использованием RAxML вер. V8.2.10 в операционной системе Linux Debian. Для анализа использовалась команда raxmlHPC-PTHREADS, и для каждого раздела была указана модель GTR + G (команда -m). Дерево ML было идентифицировано для исходной последовательности, а дополнительные деревья были определены для 2000 наборов данных с повторной выборкой бутстрапов, чтобы указать уровни поддержки ветвей (опция — #). Внешняя группа не была указана, поэтому для отображения дерево машинного обучения имело корень из средней точки.
Анализ селекции
Анализ селекции был проведен на (некорневом) дереве яДНК, которое мы вывели, и стартовый и стоп-кодоны не были включены. Мы использовали программу CODEML в PAML версии 4.8, чтобы обнаружить сигнатуры выбора по кодону, по кодону и ветви.
Свидетельство отбора по кодону
В PAML версии 4.8 (Ян 2007) правдоподобия были получены с использованием моделей сайтов M7 и M8, где соотношение несинонимичных и синонимичных замен (ω) следует бета-распределению (M7) и с дополнительным класс сайтов при положительной селекции (ω> 1) (M8). Полное описание моделей доступно в руководстве PAML (http://abacus.gene.ucl.ac.uk/software/pamlDOC.pdf). Для сравнения обеих моделей использовались тесты отношения правдоподобия (LRT). Считалось, что сайты прошли положительный отбор, когда LRT соответствующих генов были значимыми для сравнений M7-M8. Апостериорные вероятности для классов сайтов были получены с использованием байесовского эмпирического байесовского подхода (BEB) (Yang et al. 2005). Это позволило идентифицировать положительно выбранные сайты в рамках моделей M8.Мы посчитали, что существуют убедительные доказательства того, что кодирующий сайт находится под положительным отбором, когда апостериорная вероятность BEB была> 0,95 и ω> 1. Это представляет собой жесткое требование, учитывая, что ω вряд ли будет в среднем> 1 по всей группе.
Полное описание моделей доступно в руководстве PAML (http://abacus.gene.ucl.ac.uk/software/pamlDOC.pdf). Для сравнения обеих моделей использовались тесты отношения правдоподобия (LRT). Считалось, что сайты прошли положительный отбор, когда LRT соответствующих генов были значимыми для сравнений M7-M8. Апостериорные вероятности для классов сайтов были получены с использованием байесовского эмпирического байесовского подхода (BEB) (Yang et al. 2005). Это позволило идентифицировать положительно выбранные сайты в рамках моделей M8.Мы посчитали, что существуют убедительные доказательства того, что кодирующий сайт находится под положительным отбором, когда апостериорная вероятность BEB была> 0,95 и ω> 1. Это представляет собой жесткое требование, учитывая, что ω вряд ли будет в среднем> 1 по всей группе.
Селективное давление среди различных групп генов (факторы сборки и Комплексы I – V) сравнивали с использованием отношений правдоподобия отдельных генов в рамках модели сайта M7-M8.
Свидетельство выбора на кодон, на ветвь
В модели участка ветвления PAML (Yang 2007), известной как модель A, ветви разделены на группы переднего плана и группы фона. Эта модель предоставила более консервативные результаты LRT, чем результаты модели тестирования на основе ветвей. В линиях переднего плана часть сайтов находится под положительным отбором (ω> 1), тогда как фоновые линии имеют класс консервативных сайтов с ω между 0 и 1 и класс нейтральных сайтов с фиксированным ω равным 1. Поскольку Было трудно выбрать ветви переднего плана априори, мы определили каждую возможную ветвь как ветвь переднего плана, а затем проверили, присутствует ли положительный выбор.Выбор был протестирован путем сравнения двух моделей: нулевой модели A, в которой участки в линиях переднего плана установлены на ω = 1, с альтернативной моделью A, в которой ω> 1. Мы использовали метод BEB для получения апостериорных вероятностей. Мгновенное изменение скорости кодонов i на j зависит от параметра π j (равновесная частота кодона j ), который был оценен с использованием частот нуклеотидов, наблюдаемых в каждой из трех позиций кодона в гене (F3 × 4 модель).
Эта модель предоставила более консервативные результаты LRT, чем результаты модели тестирования на основе ветвей. В линиях переднего плана часть сайтов находится под положительным отбором (ω> 1), тогда как фоновые линии имеют класс консервативных сайтов с ω между 0 и 1 и класс нейтральных сайтов с фиксированным ω равным 1. Поскольку Было трудно выбрать ветви переднего плана априори, мы определили каждую возможную ветвь как ветвь переднего плана, а затем проверили, присутствует ли положительный выбор.Выбор был протестирован путем сравнения двух моделей: нулевой модели A, в которой участки в линиях переднего плана установлены на ω = 1, с альтернативной моделью A, в которой ω> 1. Мы использовали метод BEB для получения апостериорных вероятностей. Мгновенное изменение скорости кодонов i на j зависит от параметра π j (равновесная частота кодона j ), который был оценен с использованием частот нуклеотидов, наблюдаемых в каждой из трех позиций кодона в гене (F3 × 4 модель).
Корреляция между количеством кодонов при выборе и изменением высоты в ветвях
Взаимосвязь между избирательным давлением и изменениями высоты на разных ветвях количественно оценивалась частичной корреляцией между количеством положительно выбранных сайтов в каждой ветви и изменением высоты вдоль этой ветви , контролируя длину ветки. Количество положительно выбранных сайтов рассчитывали как долю положительно выбранных сайтов, умноженную на длину аминокислоты каждого гена.Высота каждого предкового узла оценивалась с помощью пакета phytools R на основе оцененного дерева (см.) И известной высоты выборки каждого образца (дополнительная таблица S2). Высота предкового узла использовалась для расчета изменения высоты вдоль каждой ветви.
Расчетная высота местообитания видов рода Phrynocephalus (цветовая шкала показывает высоту от 602 до 4541 м). Цвета на концах представляют высоту в точках отбора проб, а высота в предковых узлах была определена, как описано в разделе «Материалы и методы». Ветви пронумерованы, а звездочками обозначено 11 ветвей (№ 1, 4, 5, 6, 7, 8, 10, 11, 19, 20, 26), представляющих виды, которые эволюционировали от низко- к высокогорных местообитаний. Все виды, спускающиеся с ветви № 1, живородящие, а все виды, спускающиеся с ветви № 2, — яйцекладущие. Изменения высот дельты 28 ответвлений перечислены в дополнительной таблице S4.
Ветви пронумерованы, а звездочками обозначено 11 ветвей (№ 1, 4, 5, 6, 7, 8, 10, 11, 19, 20, 26), представляющих виды, которые эволюционировали от низко- к высокогорных местообитаний. Все виды, спускающиеся с ветви № 1, живородящие, а все виды, спускающиеся с ветви № 2, — яйцекладущие. Изменения высот дельты 28 ответвлений перечислены в дополнительной таблице S4.
Параллельная эволюция ветвей, эволюционирующих от низкого к высокому уровню
Мы также исследовали, имеют ли ветви, развивающиеся от низкого к высокому уровню, тенденцию показывать доказательства отбора на одних и тех же участках (кодоны или гены) по сравнению с линиями, развивающимися от высокого к низкому уровню. .Мы проверили отбор как на уровне генов, так и на уровне кодонов. Для этого анализа мы использовали только ветви, в которых по крайней мере 1 сайт (кодон или ген) находился под положительным отбором. Критерием того, чтобы сайт считался находящимся под положительным отбором, было то, что он имел значимые значения отношения lnL и значения омега больше 1.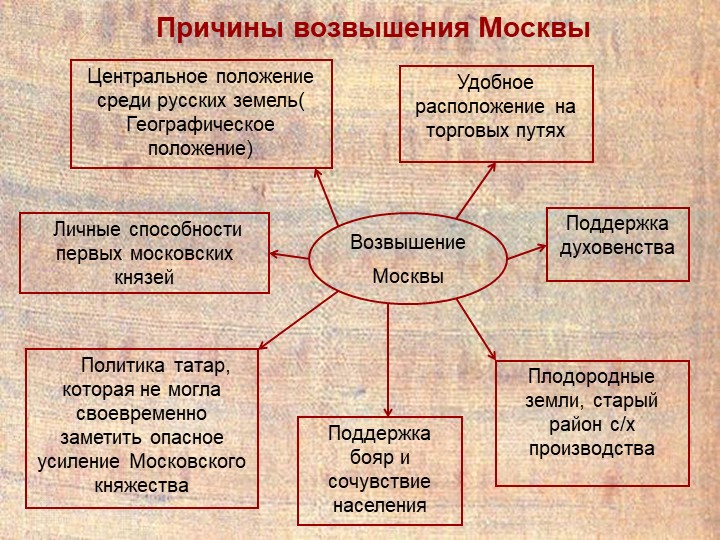 Для кодоновых сайтов мы дополнительно требовали апостериорную вероятность BEB выше 0,95. Для кодонов это включало в общей сложности 16 ветвей, в том числе 8 ветвей, эволюционирующих от низких к высоким, и 8 ветвей, эволюционирующих от высоких к низким уровням.Для сайтов генов мы включили 8 ветвей, эволюционирующих от низких к высоким, и 11 ветвей, эволюционирующих от высоких к низким уровням ().
Для кодоновых сайтов мы дополнительно требовали апостериорную вероятность BEB выше 0,95. Для кодонов это включало в общей сложности 16 ветвей, в том числе 8 ветвей, эволюционирующих от низких к высоким, и 8 ветвей, эволюционирующих от высоких к низким уровням.Для сайтов генов мы включили 8 ветвей, эволюционирующих от низких к высоким, и 11 ветвей, эволюционирующих от высоких к низким уровням ().
Мы проверили, есть ли более сильное перекрытие между выбранными участками в ветвях, которые перешли на большую высоту, чем в других ветвях. Мы называем ответвления, связанные с переходом на большую высоту (обычно для живородящих видов в QTP), «целевыми ветвями». В частности, мы проверяем гипотезу о том, что если сайт находится в режиме выбора в одной целевой ветви, то с большей вероятностью он будет выбран в других целевых ветвях, в большей степени, чем сайты, выбранные в нецелевых ветвях.
Мы представили наши данные в виде матрицы (m si ) со строками, соответствующими сайтам генов или кодонов, и столбцами, соответствующими ветвям в филогении. Записи матрицы являются двоичными, где 1 означает положительный выбор, а 0 означает отсутствие выбора. На каждом участке, s , мы берем все возможные пары целевых ветвей ( i , j ) (те, которые развиваются от низких до высоких высот) и подсчитываем, сколько пар сайт находится под положительным выбором в обеих ветвях ( m si = m sj = 1).Мы определяем тестовую статистику γ как сумму этого подсчета по всем сайтам. Формально в матричных обозначениях
Записи матрицы являются двоичными, где 1 означает положительный выбор, а 0 означает отсутствие выбора. На каждом участке, s , мы берем все возможные пары целевых ветвей ( i , j ) (те, которые развиваются от низких до высоких высот) и подсчитываем, сколько пар сайт находится под положительным выбором в обеих ветвях ( m si = m sj = 1).Мы определяем тестовую статистику γ как сумму этого подсчета по всем сайтам. Формально в матричных обозначениях
γ = # {(i, j): i и j — целевые ветви и msi = msj = 1}.
Мы сгенерировали k случайных перестановок матрицы при предположении независимости с фиксированными маржинальными значениями. Мы выполнили перестановки, используя пакет permat в R (https://www.rdocumentation.org/packages/vegan/versions/2.4-2/topics/permat). Были применены три метода перестановки (swap, tswap и quasiswap) (Patefield 1981; Hardy 2008).Мы обусловливаем маргинальные значения матрицы, чтобы учесть вариации в количестве сайтов или генов, подвергаемых отбору, среди ветвей и вариации в количестве ветвей, подвергаемых отбору, среди сайтов или генов. Для каждой из переставленных матриц вычислялась тестовая статистика γ. Затем мы сравнили наблюдаемую статистику теста с нулевым распределением, полученным для k перестановок. Односторонний тест рассчитывали как долю случаев, когда наблюдаемое значение тестовой статистики γ меньше, чем тестовая статистика γ переставленной матрицы.Мы запускали каждый алгоритм перестановки для 1 миллиона итераций со 100 000 итераций приработки.
Для каждой из переставленных матриц вычислялась тестовая статистика γ. Затем мы сравнили наблюдаемую статистику теста с нулевым распределением, полученным для k перестановок. Односторонний тест рассчитывали как долю случаев, когда наблюдаемое значение тестовой статистики γ меньше, чем тестовая статистика γ переставленной матрицы.Мы запускали каждый алгоритм перестановки для 1 миллиона итераций со 100 000 итераций приработки.
Результаты
Гены и филогенетика
После выравнивания собранных транскриптомов со 125 последовательностями генов ПКР и удаления альтернативных участков сплайсинга (как описано в материалах и методах), мы выбрали в общей сложности 106 генов, имеющих общую длину 70,341 п.н. каждый ген варьирует от 87 до 2526 п.н. Подробная информация об проанализированных генах приведена в дополнительной таблице S3.
Филогенетический анализ с использованием RAxML v8.2.X выявил 2 основные клады, представленные, соответственно, всеми живородящими или яйцекладущими видами (), что согласуется с ранее опубликованными топологиями, основанными в основном на наборах данных мтДНК (Jin and Brown 2013).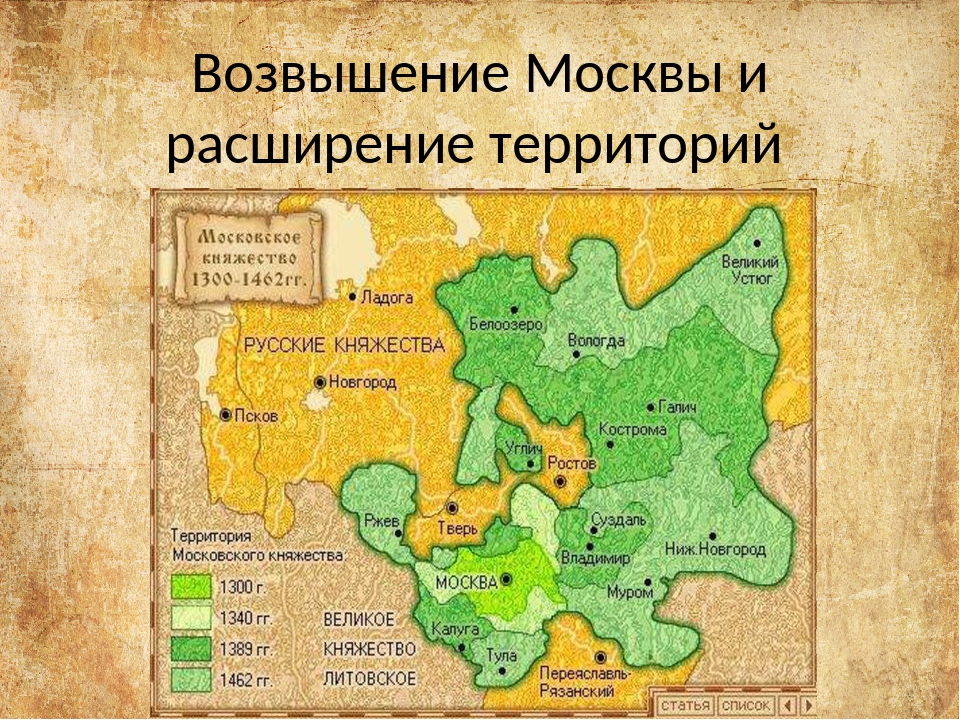 Наивысшие значения начальной загрузки (100) были получены для всех основных узлов ().
Наивысшие значения начальной загрузки (100) были получены для всех основных узлов ().
Филогенетическое дерево RAxML оценено с использованием последовательностей 106 ПКР и сложных генов факторов сборки. Клады, заштрихованные серым цветом, показывают виды из низинных областей, а другие клады обитают на возвышенностях Тибетского плато.Наивысшие значения начальной загрузки (100) были оценены для всех узлов.
Сравнение положительного отбора среди комплексов и генов
Сравнения селективного давления среди генов в различных комплексах были сделаны на основе логарифмических отношений правдоподобия (LLR) генов, оцененных с использованием модели M7-M8. Высокое отношение lnL в модели M7-M8 указывает на свидетельство отбора. Отношения логарифма правдоподобия (lnL) всех генов перечислены в дополнительной таблице S3. Используя непараметрический тест, мы обнаружили незначительную разницу в давлении положительного отбора между комплексами и комплексными факторами сборки (критерий Вилкоксона W = 1,505, P = 0. 1029,), при этом респираторный комплекс подвергается большему отбору, чем сложные факторы сборки. Однако не было обнаружено значительных различий между 5 комплексами (Крускал – Уоллис H = 3.3484, df = 4, P = 0.5013,). Мы отмечаем, что при обнаружении различий в давлении отбора среди комплексов есть оговорка, что были включены только 4 гена для Комплекса II, и это не показывает доказательств отбора. Не было обнаружено значительных различий между комплексами I, III, IV и V, когда тест был повторен, включая только эти группы (Kruskal – Wallis H = 1.2839, df = 3, P = 0,733). Пять генов с наиболее убедительными доказательствами отбора, измеренными их отношениями большого правдоподобия (> 20), — это NDUFS7 (который является субъединицей Комплекса I), COX6C (Комплекс IV), COX5B (Комплекс IV), NDUFC2 (Комплекс I), и ATP5PD (Комплекс V).
1029,), при этом респираторный комплекс подвергается большему отбору, чем сложные факторы сборки. Однако не было обнаружено значительных различий между 5 комплексами (Крускал – Уоллис H = 3.3484, df = 4, P = 0.5013,). Мы отмечаем, что при обнаружении различий в давлении отбора среди комплексов есть оговорка, что были включены только 4 гена для Комплекса II, и это не показывает доказательств отбора. Не было обнаружено значительных различий между комплексами I, III, IV и V, когда тест был повторен, включая только эти группы (Kruskal – Wallis H = 1.2839, df = 3, P = 0,733). Пять генов с наиболее убедительными доказательствами отбора, измеренными их отношениями большого правдоподобия (> 20), — это NDUFS7 (который является субъединицей Комплекса I), COX6C (Комплекс IV), COX5B (Комплекс IV), NDUFC2 (Комплекс I), и ATP5PD (Комплекс V).
LLR (lnLR) отбора в основных генах (субъединицах) ПКР и генах факторов сборки ПКР. ( A ) Доказательства положительного отбора выше для генов, составляющих субъединицы комплексов, чем для комплексных факторов сборки (тест Вилкоксона W = 1,505, P = 0.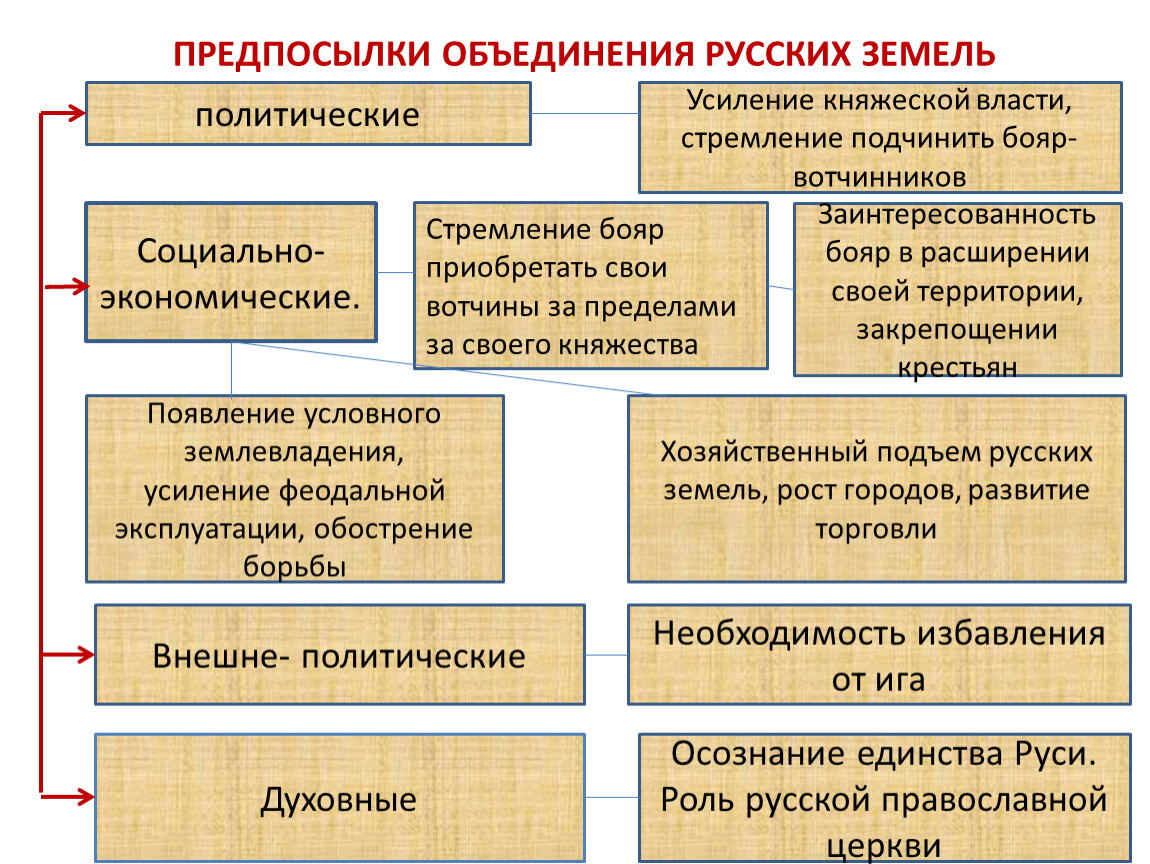 1029. ( B ) Нет свидетельств разницы в положительном отборе между разными комплексами (Краскал – Уоллис H = 3,3484, P = 0,5013).
1029. ( B ) Нет свидетельств разницы в положительном отборе между разными комплексами (Краскал – Уоллис H = 3,3484, P = 0,5013).
Положительный отбор и параллельная эволюция ветвей среди градиентов высоты
Мы обнаружили положительную корреляцию между количеством положительно выбранных кодонов и абсолютным изменением высоты среди 28 ветвей, когда средняя длина ветвей контролировалась для (частичная корреляция Пирсона: r = 0.522, P = 0,005, df = 25,). Корреляция оставалась значимой или незначительно незначимой, когда мы анализировали отдельно ветви, развивающиеся от низкого к высокому, и ветви от высокого к низкому уровню (ветви от высокого к низкому: r = −0,635, P = 0,036, df = 9, ответвления от низких к высоким отметкам: r = 0,506, P = 0,054, df = 13,). Это указывает на то, что ветви с более значительными высотными изменениями подверглись более сильному положительному отбору. Мы отмечаем, что некоторые корреляции являются маргинальными и не могут быть воспроизведены, когда сила отбора измерялась как сумма LLR отбора на всех участках в ветви (). Подробные данные, включая дельта-подъемы, пропорции кодонов положительного отбора, суммы отношений правдоподобия для 106 генов и средние длины ветвей, используемые для корреляционного анализа, были перечислены в дополнительной таблице S4.
Мы отмечаем, что некоторые корреляции являются маргинальными и не могут быть воспроизведены, когда сила отбора измерялась как сумма LLR отбора на всех участках в ветви (). Подробные данные, включая дельта-подъемы, пропорции кодонов положительного отбора, суммы отношений правдоподобия для 106 генов и средние длины ветвей, используемые для корреляционного анализа, были перечислены в дополнительной таблице S4.
Частичная корреляция между изменением высоты и силой положительного отбора с учетом влияния длины ветви.Частичная корреляция была значимой, когда сила отбора измерялась как количество кодонов при отборе ( A , B , C ), но не была значимой, когда отбор измерялся как сумма LLR тестов отбора ( D , E , F ). (A, D) Включены все ветви, использовалась абсолютная величина изменения высоты. (B, E) Включены только ветви, развивающиеся от высокого к низкому уровню (отрицательное изменение высоты). (C, F) Включены только ветви, развивающиеся от низкого до высокого уровня (положительное изменение высоты).
(C, F) Включены только ветви, развивающиеся от низкого до высокого уровня (положительное изменение высоты).
Затем мы искали сигналы параллельной эволюции между ветвями, развивающимися от низкого до высокого уровня. В частности, мы использовали тест на основе перестановок, чтобы определить, демонстрируют ли похожие кодоны или гены сигналы отбора на этих ветвях. Для этого анализа мы использовали только ветви, в которых хотя бы 1 сайт (кодон или ген) находился под положительным отбором. Матрицы кодонов и генных ветвей, используемые для анализа перестановок, перечислены в дополнительных таблицах S5 и S6 (см. Также «Методы» для получения более подробной информации о тесте перестановок).
Тесты были выполнены с использованием 3 различных методов перестановки матриц (quasiswap, swap и tswap), подтвердившие, что набор целевых ветвей, эволюционирующих от низкого к высокому уровню, разделяет больше кодонов при отборе, чем ветви, развивающиеся от высокого к низкому ( P -значения были равны нулю во всех трех методах перестановки). Мы не наблюдали этот сигнал на уровне гена (значения P были выше 0,22 для всех 3 методов перестановки).
Мы не наблюдали этот сигнал на уровне гена (значения P были выше 0,22 для всех 3 методов перестановки).
Среди кодонов, подвергшихся положительному отбору в более чем одной ветви, эволюционирующей от низкого до высокого уровня, было обнаружено только одно повторяющееся изменение состояния аминокислоты, изменение с «Ala» на «Gly» в гене NDUFS7 (дополнительная таблица S7).
Обсуждение
В этом исследовании изучалась селекция по большинству известных ядерных генов, составляющих ПКР, а также по сложным факторам сборки. Мы показали, что нет существенной разницы в селективном давлении между комплексами, а также между комплексными субъединицами и факторами сборки. Следовательно, наши результаты не подтверждают предыдущие исследования (Mishmar et al.2006; Yu et al.2011; Jin et al.2018) с использованием только митохондриальных генов, которые обнаружили более высокое давление отбора на Комплекс I, чем на другие комплексы Phrynocephalus .Наше исследование согласуется с предыдущими исследованиями, которые продемонстрировали доказательства положительного отбора и связанных с климатом различий в последовательностях и экспрессии генов ПКР у ряда видов животных с широкими биогеографическими диапазонами (Toews et al. 2014; Garvin et al. 2015; Моралес и др., 2018).
2014; Garvin et al. 2015; Моралес и др., 2018).
Результаты частичного корреляционного анализа показали, что ветви, развивающиеся при больших изменениях высоты, как правило, испытывают более сильное давление отбора, когда давление отбора измеряется как количество положительно выбранных кодонов.Другой показатель силы выбора по ветви — это сумма LLR, сравнивающих модели с (M8) и без выбора (M7). Мы не обнаружили значимой корреляции между силой отбора, измеренной суммой LLR, и изменением высоты вдоль ветви. Однако наши результаты поднимают интересную гипотезу о том, что виды, которые претерпели более значительные высотные изменения среды обитания в своей эволюционной истории, испытывают более высокий уровень избирательного давления на гены ПКР. Эта гипотеза может быть дополнительно проверена с дополнительными данными по Phrynocephalus или другим видам, живущим в чрезвычайно высоких средах, а также по другим генам, которые могут находиться под отбором из-за изменения высоты.
Наш анализ перестановок ветвей кодонов показывает, что ветви, представляющие эволюцию от малой высоты к большой, имели более частый отбор по одним и тем же кодонам, чем ветви, возникающие от большой высоты к низкой. Phrynocephalus произошел из низменностей Центральной Азии, и живородящий субклад (см.) Возник в QTP вместе с поднятием QTP 10 миллионов лет назад. Наши результаты согласуются с идеей, что, поскольку Phrynocephalus ящериц занимали большие высоты, эта новая среда наложила отбор на аналогичные кодоны ПКР.
Мы обнаружили только один случай параллельной эволюции аминокислотного состояния в гене NDUFS7. Этот ген по-разному экспрессируется у пищух на разных высотах в Гималаях, что позволяет предположить его роль в адаптации к высоте у других групп животных (Solari et al. 2018). Мы предполагаем, что параллельная эволюция может быть редкой из-за простой случайности, то есть, если определенный аллель не был доступен ни из общей наследственной вариации, ни из повторяющейся мутации. Это также может быть редкостью из-за компромиссов с окружающей средой, которые могут привести к тому, что аллель будет благоприятным в одной из высокогорных сред, но не в другой.Также возможно, что параллельная эволюция на уровне кодонов редка, потому что функциональная конвергенция может происходить при отборе в разных генах или кодонах.
Это также может быть редкостью из-за компромиссов с окружающей средой, которые могут привести к тому, что аллель будет благоприятным в одной из высокогорных сред, но не в другой.Также возможно, что параллельная эволюция на уровне кодонов редка, потому что функциональная конвергенция может происходить при отборе в разных генах или кодонах.
Пониманию генетической адаптации животных, живущих в чрезвычайно высокогорных средах QTP, в последнее время уделяется все больше внимания (Sun et al. 2018). Здесь мы проанализировали большинство ядерных генов ПКР и показали доказательства того, что виды, живущие на больших высотах, могут подвергаться более сильному положительному отбору, влияющему на дыхательную цепь, чем близкородственные виды, живущие на более низких высотах.Однако функциональные последствия генетической адаптации ПКР, показанные в этом исследовании, остаются непроверенными. Хотя исследования метаболизма рептилий, обитающих на большой высоте, остаются ограниченными, недавнее исследование показало более низкую частоту дыхания и более высокую эффективность производства АТФ митохондриями у P. erythrurus из высокогорных областей QTP, чем у P. przewalskii из низкие высоты за пределами QTP, когда оба вида содержались при давлении кислорода, аналогичном таковому в их соответствующих средах обитания (Tang et al.2013). Это могут быть функциональные последствия положительно выбранных изменений кодонов, обнаруженных в этом исследовании.
erythrurus из высокогорных областей QTP, чем у P. przewalskii из низкие высоты за пределами QTP, когда оба вида содержались при давлении кислорода, аналогичном таковому в их соответствующих средах обитания (Tang et al.2013). Это могут быть функциональные последствия положительно выбранных изменений кодонов, обнаруженных в этом исследовании.
Таким образом, наше исследование показывает, что повышение приводит к важным различиям в выборе митохондриальных ПКР Phrynocephalus ящериц с Тибетского плато. Группы, колонизировавшие высоко с более низких высот, по-видимому, испытали более сильные селективные силы, чем те, которые двигались в противоположном направлении. Повышенный выбор, кажется, связан с большим сдвигом по высоте.Наконец, давление отбора одинаково для генов, кодирующих субъединицы ПКР и факторов сборки ПКР.
Дополнительные материалы
zoaa056_Supplementary_Data
Благодарности
Мы хотим поблагодарить Кайлонга Чжана за помощь в сборе образцов. Мы особенно благодарны Расмусу Нильсену из Калифорнийского университета в Беркли за его любезные предложения по анализу этой работы.
Мы особенно благодарны Расмусу Нильсену из Калифорнийского университета в Беркли за его любезные предложения по анализу этой работы.
Финансирование
Работа выполнена при поддержке Национального фонда естественных наук Китая (грант №31772447) и Вторая программа научных экспедиций и исследований на Тибетское плато (STEP), грант № 2019 QZKK0501. ПРОТИВ. исследование проводилось в рамках Программы фундаментальных исследований НИУ ВШЭ.
Заявление о доступности данных
Данные о географических точках отбора проб и всех измеренных цветах спины и субстрата хранятся в FigShare (doi: 10.6084 / m9.figshare.10308572). Соответствующие авторы несут ответственность за любые личные требования к материалам.
Список литературы
- Акунья-Кастровьехо Д., Мартин М., Масиас М., Эскамес Дж., Леон Дж.и др., 2001. Мелатонин, митохондрии и клеточная биоэнергетика. J. Pineal Res 30: 65–74. [PubMed] [Google Scholar]
- Цай Цюй, Цянь Х, Ланг И, Ло И, Сюй Дж.
 и др., 2013.
Последовательность генома наземной синицы pseudopodoces humilis и ее адаптация к большой высоте. Геном Биол 14: R29. [Бесплатная статья PMC] [PubMed] [Google Scholar]
и др., 2013.
Последовательность генома наземной синицы pseudopodoces humilis и ее адаптация к большой высоте. Геном Биол 14: R29. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Камю М.Ф., Вольф Дж. Н., Сгро К. М., Даулинг Д. К., 2017. Экспериментальное подтверждение того, что естественный отбор сформировал широтное распределение митохондриальных гаплотипов у австралийской Drosophila melanogaster .Mol Biol Evol 34: 2600–2612. [PubMed] [Google Scholar]
- Fiedorczuk K, Letts J, Degliesposti G, Kaszuba K, Skehel M. и др., 2016. Атомная структура всего митохондриального комплекса млекопитающих I. Nature 538: 406–410. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Fonseca RRD, Johnson WE, Obrien SJ, Ramos MJ, Antunes A, 2008. Адаптивная эволюция митохондриального генома млекопитающих. BMC Genom 9: 119–119. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Фут А.Д., Морин П.А., Дурбан Д.В., Питман Р.Л., Уэйд ПР.и др., 2011.
Положительный отбор на митогеном косатки.
 Biol Lett 7: 116–118. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Biol Lett 7: 116–118. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Гарвин М.Р., Торгаард Г.Х., Нарум С.Р., 2015. Дифференциальная экспрессия генов, контролирующих дыхание, способствует термической адаптации краснозобой форели ( Oncorhynchus mykiss gairdneri ). Genome Biol Evol 7: 1404–1414. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Грабхерр М., Хаас Б.Дж., Яссур М., Левин Дж.З., Томпсон Д.А. и др., 2011.Сборка полноразмерного транскриптома из данных РНК-seq без референсного генома. Nat Biotechnol 29: 644–652. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Харди О.Дж., 2008. Тестирование пространственной филогенетической структуры местных сообществ: статистические характеристики различных нулевых моделей и тестовая статистика для локально нейтрального сообщества. Дж. Экол 96: 914–926. [Google Scholar]
- Хассанин А., Ропике А., Кулу А., Круод С., 2009.
Эволюция митохондриального генома у млекопитающих, живущих на большой высоте: новые выводы из исследования трибы Caprini (Bovidae, Antilopinae).
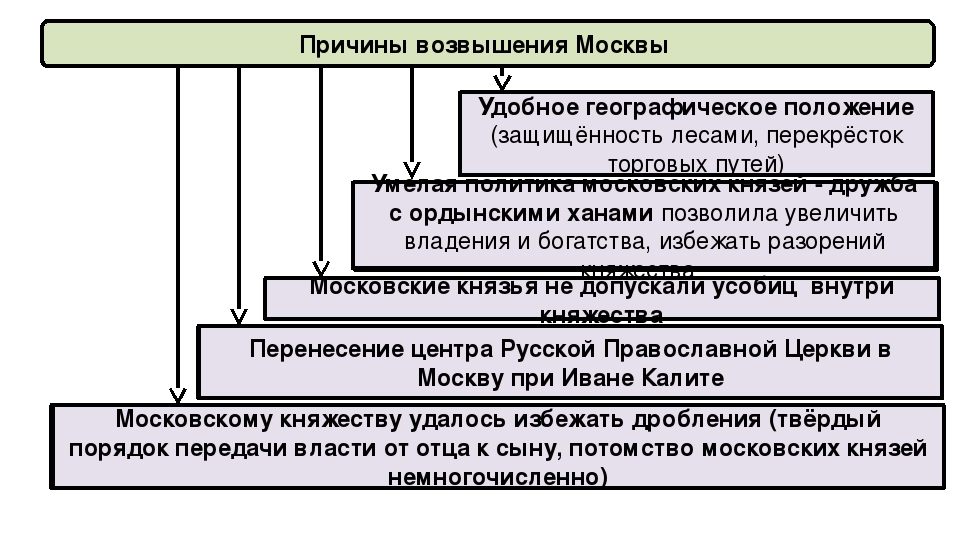 J Mol Evol 68: 293–310. [PubMed] [Google Scholar]
J Mol Evol 68: 293–310. [PubMed] [Google Scholar] - Хейс Дж., О’Коннор К.С., 1999. Естественный отбор по термогенной способности высотных мышей-оленей. Evolution 53: 1280–1287. [PubMed] [Google Scholar]
- Джин Ю.Т., Браун Р.П., 2013. История видов и время расхождения живородящих и яйцекладущих ящериц с головой китайской жабы Phrynocephalus на Цинхай-Тибетском плато. Mol Phylogenet Evol 68: 259–268. [PubMed] [Google Scholar]
- Цзинь Ю.Т., Браун Р.П., Лю Н.Ф., 2008.Кладогенез и филогеография ящерицы Phrynocephalus vlangalii (agamidae) на Тибетском плато. Мол Экол 17: 1971–1982. [PubMed] [Google Scholar]
- Цзинь Ю.Т., Ли Дж.К., Лю Н.Ф., 2013. Связанные с высотами различия в жизненных чертах среди линий Phrynocephalus на тибетском плато: следуют ли они типичным чешуйчатым экогеографическим образцам ?. Дж. Зоол 290: 293–301. [Google Scholar]
- Джин И, Ву И, Тонг Х, Сонг С, Чжан Л.
и др., 2018.
Эволюционный анализ митохондриально кодируемых белков ящериц с головой жабы Phrynocephalus по высотному градиенту.
 BMC Genom 19: 185. [Бесплатная статья PMC] [PubMed] [Google Scholar]
BMC Genom 19: 185. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Лашанс Дж, Тишкофф С.А., 2013. Популяционная геномика адаптации человека. Annu Rev Ecol Evol Syst 44: 123–143. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Letts J, Fiedorczuk K, Sazanov L, 2016. Архитектура респираторных суперкомплексов. Природа 537: 644–648. [PubMed] [Google Scholar]
- Ли И, Рен Зи, Шедлок А.М., Ву Дж., Санг Л. и др., 2013. Высотная адаптация рыб-шизоторацинов (Cyprinidae), выявленная анализом митохондриального генома.Ген 517: 169–178. [PubMed] [Google Scholar]
- Лим MCW, Витт CC, Грэм CH, Давалос Л.М., 2019. Параллельная молекулярная эволюция путей, генов и участков у высокогорных колибри, выявленная с помощью сравнительной транскриптомики. Genome Biol Evol 11: 1573–1585. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Лобо-Ярне Т., Угальде С., 2018.
Суперкомплексы дыхательной цепи: строение, функции и биогенез. Semin Cell Dev Biol 76: 179–190.
 [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar] - Ло И, Чен И, Лю Ф, Юци Г., 2003.Митохондриальный геном тибетского дикого осла Equus kiang обнаруживает замены в nadh, которые могут отражать эволюционную адаптацию к холоду и гипоксии. Asia Life Sci 21: 1–11. [Google Scholar]
- Мишмар Д., Руис-Эсини Э, Мондрагон-Паломино М., Прокаччо V, Гаута Б. и др., 2006. Адаптивный отбор субъединиц I митохондриального комплекса при облучении приматов. Ген 378: 11–18. [PubMed] [Google Scholar]
- Моралес Х.Е., Павлова А., Амос Н., Майор Р., Килиан А. и другие.2018. Согласованное расхождение митогеномов и кластера митонуклеарных генов в линиях птиц, населяющих разный климат. Природа, экология и эволюция 2: 1258–1267. DOI: 10.1038 / s41559-018-0606-3. [PubMed] [CrossRef] [Google Scholar]
- Нгатиа Дж. Н., Лан Т. Д., Динь Т. Д., Чжан Л., Ахмед А. К..
и др., 2019.
Сигналы положительного отбора в митохондриальных генах, кодирующих белок шерстистого мамонта: адаптация к экстремальным условиям ?.
 Ecol Evol 9: 6821–6832. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Ecol Evol 9: 6821–6832. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Пейтфилд WM, 1981.Алгоритм as 159: эффективный метод создания случайных таблиц размера r × c с заданными суммами строк и столбцов. J R Stat Soc 30: 91–97. [Google Scholar]
- Резенде Э.Л., Dinizfilho JAF, 2012. Филогенетический анализ: сравнение видов для вывода адаптаций и физиологических механизмов. Compr Physiol 2: 639–674. [PubMed] [Google Scholar]
- Шмидт Т.Р., Ву В., Гудман М., Гроссман Л.И., 2001. Эволюция взаимодействия кодируемых ядер и митохондрий субъединиц в цитохром с оксидазе. Mol Biol Evol 18: 563–569.[PubMed] [Google Scholar]
- Солари К., Рамакришнан У., Хадли Э.А., 2018. Экспрессия генов вовлечена в способность пищух занимать гималайский градиент высоты. PLoS ONE 13: e0207936. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Соловьева Е.Н., Лебедев В.С., Дунаев Е.А., Назаров Р.А., Банникова А.А.
и др., 2018.
Аридизация кайнозоя в Центральной Евразии сформировала разнообразие жабоголовых агам ( Phrynocephalus ; Agamidae, Reptilia).
 PeerJ 6: e4543. [Бесплатная статья PMC] [PubMed] [Google Scholar]
PeerJ 6: e4543. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Сунь И, Фу Т, Джин Дж, Мерфи Р.У., Хиллис Д.М.и др., 2018. Группы видов, распределенные по высотным градиентам, демонстрируют конвергентную и непрерывную генетическую адаптацию к высоким высотам. Proc Natl Acad Sci USA 115: e10634 – e10641. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Тан X, Синь Y, Ван Х, Ли В, Чжан Ю. и др., 2013. Метаболические характеристики и реакция на большую высоту в Phrynocephalus erythrurus (Lacertilia: Agamidae), ящерица, обитающая на высоте выше, чем любые другие живущие ящерицы в мире.PLoS ONE 8: e71976. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Тэйвс ДПЛ, Мандич М., Ричардс Дж. Г., Ирвин Д.Е., 2014. Миграция, митохондрии и камышевка. Evolution 68: 241–255. [PubMed] [Google Scholar]
- Ян З, 2007. Память 4: филогенетический анализ методом максимального правдоподобия. Mol Biol Evol 24: 1586–1591. [PubMed] [Google Scholar]
- Ян З.
 , Вонг WSW, Нильсен Р., 2005.
Байесовский эмпирический байесовский вывод аминокислотных участков при положительном отборе. Mol Biol Evol 22: 1107–1118.[PubMed] [Google Scholar]
, Вонг WSW, Нильсен Р., 2005.
Байесовский эмпирический байесовский вывод аминокислотных участков при положительном отборе. Mol Biol Evol 22: 1107–1118.[PubMed] [Google Scholar] - Ю Л, Ван Х, Тинг Н, Чжан И, 2011. Митогеномный анализ китайских курносых обезьян: свидетельство положительного отбора генов наддегидрогеназы в условиях высотной адаптации. Митохондрия 11: 497–503. [PubMed] [Google Scholar]
- Юань М.Л., Чжан К.Л., Чжан Л., Цзя К.Л., Ли ХР. и др., 2018. Филогения митохондрий, история дивергенции и высотная адаптация пастбищных гусениц (Lepidoptera: Lymantriinae: Gynaephora ), населяющих Тибетское плато.Мол Филогенет Эвол 122: 116–124. [PubMed] [Google Scholar]
- Чжао К.Т., 1999. Phrynocephalus (Agamidae). В: Чжао Е.М., Чжао К.Т., Чжоу К.Ю., редакторы. Фауна Sinica, Reptilia (Squamata: Lacertilia). Vol. 2. Пекин: Science Press. 151–193. [Google Scholar]
- Чжоу Т., Шен Х, Ирвин Д.М., Шен Й, Чжан И, 2014.
Митогеномный анализ предполагает положительный отбор митохондриальных генов для высотной адаптации галлиформных птиц.
 Митохондрия 18: 70–75. [PubMed] [Google Scholar]
Митохондрия 18: 70–75. [PubMed] [Google Scholar] - Цинк Р.М., 2005.Естественный отбор на митохондриальной ДНК in parus и его актуальность для филогеографических исследований. Proc Biol Sci 272: 71–78. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Мехико 1968 — Спортсмены, медали и результаты
Драма и красота
Игры были благословлены многими выдающимися героинями, но не более, чем Верой Чаславской. Привлекательная и жизнерадостная чешская гимнастка завоевала четыре золотых и две серебряные медали. Ее победы были драматичными: победа над советскими гимнастками через два месяца после советского вторжения на ее родину.
НОК: 112
Спортсмены: 5,516 (781 женщины, 4435 мужчины)
События: 172
Волонтеры: Н / Д
СМИ: Н / Д
Немецкий Спортсмены
Спортсмены из ГДР соревновались в своей команде под названием «Восточная Германия». И только в 1972 году они приняли участие под официальным названием своей страны «Германская Демократическая Республика».
И только в 1972 году они приняли участие под официальным названием своей страны «Германская Демократическая Республика».
Маршрут эстафеты Олимпийского огня пошел по стопам Христофора Колумба, проследив маршрут его первого путешествия из Испании в Новый Свет.
Особый год
1968 год был чрезвычайно важным с политической точки зрения. Китайская Народная Республика оказалась в эпицентре культурной революции. Попытка либерализовать Чехословакию была подавлена советскими войсками. Французское правительство было захвачено студенческими демонстрациями, и по всей территории Соединенных Штатов проходили демонстрации за мир и гражданские права. Мексика не была забыта на этой волне революции: студенты и учителя бастовали и проводили массовые митинги протеста, кроваво подавленные на Площади трех культур.
«Сила черных»
Помимо завоеванных медалей, чернокожие американские спортсмены сделали себе имя актом расового протеста. Во время церемонии вручения медалей Томми Смит и Джон Карлос, завоевавшие золотые и бронзовые медали в беге на 200 метров, подняли кулак в черной перчатке и склонили головы, когда звучал национальный гимн своей страны. При этом они протестовали против расовой сегрегации в Соединенных Штатах и впоследствии были изгнаны из Олимпийской деревни.
При этом они протестовали против расовой сегрегации в Соединенных Штатах и впоследствии были изгнаны из Олимпийской деревни.
Внешний вид тартана
На легкой атлетике впервые был использован синтетический материал (тартан).
Официальное время
Соревнования по легкой атлетике, велоспорту, гребле, каноэ, плаванию и конному спорту были рассчитаны вручную и электронно. Впервые электронное время стало официальным.
Церемонии
12 октября 1968 года, Мексика, церемония открытия. Энрикета Базилио де Сотело (Мексика) поднимается с олимпийским факелом.
Официальное открытие Игр:
Президент Густаво Диас Ордас
Зажигание Олимпийского огня:
Норма Энрикета Базилио де Сотело (легкая атлетика)
Олимпийская присяга:
Пабло Луго Гарридо (легкая атлетика
) Присяга:Присяга официальных лиц на летних Олимпийских играх впервые была принесена в 1972 году в Мюнхене.
Самые высокие горные вершины мира
Самые высокие вершины
Горы издавна были источником трепета и вдохновения — чем выше, тем лучше. В древности они считались домами богов. В последнее время они вдохновили на создание национальных парков. И сегодня альпинисты пересекают земной шар, чтобы покорить самые высокие точки на Земле.
В древности они считались домами богов. В последнее время они вдохновили на создание национальных парков. И сегодня альпинисты пересекают земной шар, чтобы покорить самые высокие точки на Земле.
Большинство самых высоких гор в мире расположены в Гималайском хребте, недалеко от автономного региона Тибет.Гора Эверест, самая высокая гора в мире, находится в этом хребте. Гора Эверест также известна под своим тибетским названием Джомолунгма.
Вторая по высоте гора в мире, К2, является единственным исключением, она расположена в близлежащем горном массиве Каракорум. Эти два горных хребта в Азии охватывают все восьмитысячники, избранную группу гор, самые высокие вершины которых находятся на высоте более 8000 метров над уровнем моря.
См. названных вершин в США на высоте более 14000 футов над уровнем моря для U.С. Пикс.
Или, если вы интересуетесь фотографией природы, ознакомьтесь с нашими лучшими камерами для дронов 2020 года.
См. Также Восхождение на 14 8000-метровых вершин мира, Семь вершин, Смертные на горе Олимп: A История восхождения на Эверест и Альманах Эвереста.
| Горная вершина | Диапазон | Местоположение | Высота | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| футов | м | |||||||||||||||
| Эверест 1 | Гималаи 978197835 | 1978K2 (Годвин Остин) | Каракорам | Пакистан / Китай | 28 250 | 8,611 | ||||||||||
| Канченджанга | Гималаи | Индия / Непал | ||||||||||||||
| 27,940 | 8,516 | |||||||||||||||
| Макалу I | Гималаи | Непал / Тибет | 27,766 | 8,463 | ||||||||||||
| Дхаулагири I | Гималаи | с. ш. epal ш. epal | 26,795 | 8,167 | ||||||||||||
| Manaslu I | Гималаи | Непал | 26,781 | 8,163 | ||||||||||||
| Nanga Parbat | ||||||||||||||||
| Nanga Parbat | I975Гималаи | Непал | 26,545 | 8,091 | ||||||||||||
| Гашербрум I | Каракорам | Пакистан / Китай | 26,470 | 8,068 | 8,047||||||||||||
| Гашербрум II | Каракорам | Пакистан / Китай | 26,360 | 8,035 | ||||||||||||
| Шишма Пангма (Госинтхан) | Гималаи | Гималаи | ,21975Гималаи Гималаи | Непал | 26041 | 7937 | ||||||||||
| Гьячунг-Канг | Гималаи | Непал | 25910 | 7897 | ||||||||||||
| Дастогхил | Каракорум | Пакистан | 25858 | 7882 | ||||||||||||
| Хималчули | Гималаи | Непал | 25801 | 7864 | ||||||||||||
| Nuptse | Гималаи | Непал | 25726 | 7841 | ||||||||||||
| Нанда Деви | Гималаи | Индия | 25663 | 7824 | ||||||||||||
| Машербрум | Каракорум | Кашмир 2 | 25,660 | 7,821 | ||||||||||||
| Ракапоши | Каракорам | Пакистан | 25,551 | 25,551 | 25,551 | Пакистан | 25,551 | 1983 25,461 | 7,761 | |||||||
| Камет | Гималаи | Индия / Тибет | 25,446 | 7,756 | ||||||||||||
| Намча Барва | ||||||||||||||||
| Гималаи Гималаи | Тибет | 25355 | 7728 | |||||||||||||
| Улугмузтаг | Куньлунь | Тибет | 25340 | 7723 | ||||||||||||
| Кунгур | Музтагата | Китай | 25325 | 7719 | ||||||||||||
| Тирич Мир | Гиндукуш | Пакистан | 25,230 | 7,690 | ||||||||||||
| Сасер Кангри | Каракорам | Индия | 25,172 | 7678 | II 7 657 | |||||||||||
| Минья Конка (Гонгга Шан) | Даксуэ Шан | Китай | 24900 | 7,590 | ||||||||||||
| Кула-Кангри | Гималаи | 96778 | Тибет | 24780 | 7553 | |||||||||||
| Музтагата | Музтагата | Китай | 24757 | 7546 | ||||||||||||
| Skyang Kangri | Гималаи | Kashmir | 24750 | 7544 | ||||||||||||
| Пик Исмаила Самани (ранее Пик Коммунизма) | Памир | Таджикистан | 24,590 | 7,495 | ||||||||||||
| Пик Чонсон | Гималаи | Непал | ,478919beda | Кыргызстан | 24,4 06 | 7,439 | ||||||||||
| Sia Kangri | Гималаи | Кашмир | 24,350 | 7,422 | ||||||||||||
| Пик Харамош | ,3978 | Пакистан | 24,240 | 7,388 | ||||||||||||
| Пик Палатка | Гималаи | Непал | 24,165 | 7,365 | ||||||||||||
| Чомо78 | Гималайс||||||||||||||||
| Чамланг | Гималаи | Непал | 24012 | 7319 | ||||||||||||
| Kabru | Гималаи | Непал | 24002 | 7316 | ||||||||||||
| Alung Гангрий | Гималаи | Тибет | 24000 | 7315 | ||||||||||||
| Балторо Кангри | Гималаи | Кашмир | 23,990 | 7,312 | ||||||||||||
| Музтаг Ата (К-5) | Кунлун | Китай | Китай | 23860 | 7273 | |||||||||||
| Барунтз | Гималаев | Непала | 23688 | 7220 | ||||||||||||
| Непала Пик | Гималаев | Непал | 23500 | 7163 | ||||||||||||
| Amne Машены | Куньлунь | Китай | 23,490 | 7,160 | ||||||||||||
| Гаури Санкар | Гималаи | Непал / Тибет | 23,440 | 7145 | Нункун | Гимала Яс | Кашмир | 23,410 | 7,135 | |||||||
| Пик Ленина | Памир | Таджикистан / Киргизия | 23,405 | 7,134 | ||||||||||||
| Api | Гималаи | Непал | 23,399 | 7,132 | ||||||||||||
| Пауунри | Гималаи | Индия / Китай | 23,385 | ,3789198 9198 | ||||||||||||
| Korzhenevski Пик | Памир Таджикистан | 23310 | 7105 | |||||||||||||
| Kangto Гималаи | Тибет 23260 | 7090 | ||||||||||||||
| Nyainqentanglha Nyainqentanglha Shan | Китай | 23,255 | 7,088 | |||||||||||||
| Трисули | Гималаи | Индия | 23,210 | 7,074 | ||||||||||||
| Дунагири | Гималаи | Таджикистан | 22,880 | 6,974 | ||||||||||||
| Аконкагуа | Анды | Аргентина | 22,834 | 6,960 | ||||||||||||
| Охос-дель-Саладо | Бонете | Анды | Аргентина / Чили | 22,546 | 6,872 | |||||||||||
| Ама Даблам | Гималаи | Непал | 22,494 | 6,8781978 | 22,494 | 6,8671978 | 22,494 | 6,8671978 | 9 1983 6,800||||||||
| Пик Москва | Памир | Таджикистан | 22,260 | 6,785 | ||||||||||||
| Писсис | Анды | Аргентина | 22,241 | 22,241 | 22,211 | 6,770 | ||||||||||
| Уаскарн | Анды | Перу | 22,205 | 6,768 | ||||||||||||
| Llullaillaco | ||||||||||||||||
| Аргентина Анды | Аргентина | 22047 | 6720 | |||||||||||||
| Качи | Анды | Аргентина | 22,047 | 6,720 | ||||||||||||
| 6,720 | ||||||||||||||||
| Кайлас | 75Гималай | Гималай | Анды | Аргентина / Чили | 21,720 | 6,620 | ||||||||||
| Йерупаджа | Анды | Перу | 21709 | 6,617 | ||||||||||||
| Галан | Анды | Аргентина | 21 654 | 6 600 | ||||||||||||
| Эль-Муэрто | Анды | Аргентина / Чили | 21463 | 6,578 | ||||||||||||
| 6,520 | ||||||||||||||||
| Насимиенто | Анды | Аргентина | 21,302 | 6,493 | ||||||||||||
| Иллампу | Анды | |||||||||||||||
| Боливия | ,478211983 211983 Боливия | ,478211983 211983 919 78 | 6,462 | |||||||||||||
| Coropuna | Анды | Перу | 21,083 | 6,426 | ||||||||||||
| Лаудо | Анды | ,478 | 20,958 | 6,388 | ||||||||||||
| Куско | Анды | Перу | 20,945 | 6,384 | ||||||||||||
(Аусангате)
1. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
