
aforizm (Афоризмы из комедии Д.И.Фонвизина Недоросль) — документ
«Всякая вина виновата», Тарас Скотинин (действие 1 явление 4)
«Дела не делай, от дела не бегай» (Цыфиркин действие 3 явление 6),
«Дал мне Бог ученичка, боярского сынка» (Цыфиркин действие 3, явление 6),
«виной залетишь за тридевять земель, за тридесято царство» (Правдин, явление 8, действие 4)
«Суженого конём не объедешь» (Скотинин, явление 3, действие 2)
«С великим просвещение можно быть мелкому скареду1» (Стародум, явление 1 действие 3),
«На своё счастье грех пенять» (Скотинин, явление 3, действие 2)
«Век живи, век учись» (Г-жа Простакова, явление 6 действие 2)
«Ученье — вздор» (Скотинин, явление 8, действие 4)
«Где гнев, тут и милость» (Г-жа Простакова, явление 9, действие 4)
«дай мне сроку, хотя на три дня» (Г-жа Простакова, явление 5, действие 5)
«Повинную голову меч не сечёт» (Г-жа Простакова, явление 4, действие 5)
«В большом свете водятся премелкие души» (Стародум, явление 1, действие 3)
«Честнее быть без вины обойдёну, нежели без заслуг пожаловану» (Стародум, явление 1, действие 3)
«Без вины виноват» (Простаков, явление 3, действие 5)
«Без знатных дел знатное состояние ничто» (Стародум, явление 2, действие 4)
«Собака лает, ветер носит» (Цыфиркин, явление 7 действие 3)
«Угнетать рабством себе подобных беззаконно» (Стародум, явление 1, действие 5)
«Сон в руку» (Простаков, явление 4, действие 1)
«Концы в воду» (Скотинин, явление 5, действие 1)
«Видали виды» (Простаков, явление 8, действие 1)
«Белены объелся» (Митрофан, явление 4, действие 2)
«Поминай, как звали» (Митрофан, явление 6, действие 2)
«С его кулаков да за часослов» (Митрофан, явление 6, действие 2)
«Весёлым пирком, да за свадебку» (Скотинин, явление 5, действие 3)
«По добру, по здорову» (Скотинин, явление 4, действие 5)
«Наличные деньги — не наличные достоинства» (Стародум, явление 2, действие 3)
«Ему шабаш из-за доски, а меня рады в толчки» (Цыфиркин, явление 6, действие 3)
Интересно противопоставлены афоризмы и пословицы в структуре комедии Д. И. Фонвизина «Недоросль» (это отметил в своём анализе речи персонажей литературовед К.Пигарёв). Речь отрицательных персонажей (Простаковой, Скотинина) часто неграмотна и груба, но в ней встречаются народные выражения и пословицы. А положительные персонажи со своей культурной, книжной, построенной на французский манер речью пословиц почти не употребляют — зато, скажем, Стародум так и сыплет сентенциями.
И. Фонвизина «Недоросль» (это отметил в своём анализе речи персонажей литературовед К.Пигарёв). Речь отрицательных персонажей (Простаковой, Скотинина) часто неграмотна и груба, но в ней встречаются народные выражения и пословицы. А положительные персонажи со своей культурной, книжной, построенной на французский манер речью пословиц почти не употребляют — зато, скажем, Стародум так и сыплет сентенциями.
1 Скаред — скряга, скупец
Rust bridesmaid dress
Rust bridesmaid dressСуеверия и обычаи, связанные с Новым годом
19 октября, 2021Наступление Нового года отмечается давно, однако за всю многовековую историю дата, на которую приходится празднование,… >>>Книги для студентов
26 января, 2021Миру-мир, студентам — книга! Общество мертвых поэтовН. Г. КлейнбаумУченики престижной американской академии живут по определенным… >>>Главные книжные новинки ноября от Эксмо
8 ноября, 2020Новинки ноября Следующий год в ГаванеШанель КлитонНовый роман от Шанель Клитон, автора бестселлеров New York. .. >>>
.. >>>Книги на все времена
11 октября, 2020Анатомические поезда Томас Майерс Мировой научный бестселлер и настоящая библия интегративной медицины от мануального терапевта… >>>
“Пейзаж, нарисованный чаем”, Милорад Павич
25 октября, 2021″Вопрос простой: какое время видит человек, если через зеркало смотрит на часы, висящие в комнате?»… >>>Как привить любовь к чтению?
25 августа, 2021Ученые и педагоги бьют тревогу: как привить любовь к чтению у ребенка, ведь книгу заменили… >>>“Зеленый шатер”, Людмила Улицкая
12 января, 2021Оказывается, я и забыла, какой это кайф — читать настоящего писателя. Читать и думать, перечитывать, смотреть… >>>5 занимательных фактов о сериале «Чернобыль»
3 ноября, 2020Катастрофа огромного масштаба на Чернобыльской АЭС, произошедшая в 1986 году, стала основой для сериала «Чернобыль»,… >>>
“Огненный ангел”, краткое содержание
5 ноября, 2021Рупрехт встретил Ренату весной 1534 г. , возвращаясь после десяти лет службы ландскнехтом в Европе и… >>>
, возвращаясь после десяти лет службы ландскнехтом в Европе и… >>>“Грозовой перевал”, краткое содержание
5 февраля, 2021Ощутив настоятельную необходимость отдохнуть от суеты лондонского света и модных курортов, мистер Локвуд решил на… >>>“Сестра Кэрри”, краткое содержание
16 декабря, 2020В 1889 г. восемнадцатилетняя Каролина Мибер (сестра Кэрри, как её ласково называют в семье) уезжает… >>>“Страстная неделя”, краткое содержание
9 ноября, 2020Действие происходит с 19 по 26 марта 1815 г. во Франции, в течение последней перед… >>>
Как взбодриться в дождливую погоду, чтобы не унывать?
2 декабря, 2021За окном серое небо, по карнизу стучат дождевые капли, стекло покрыто водяными струйками, солнца не… >>>Лучшие книги о фитнесе
11 октября, 2021Хорошая книга принесет пользу для всех без исключения людей – занимающихся индивидуально или с инструктором. … >>>
… >>>Как помочь ребенку приспособиться к коллективу?
27 марта, 2021Когда ребенок появляется в коллективе, то он испытывает стресс, например, впервые приходит в детский сад,… >>>Виды и жанры художественной литературы
7 января, 2021История литературы занимается записью и исследованием соответствующих событий в контексте литературы (книжные публикации, изменения рынка… >>>
Баратынский Е.А. Перстень. Скачать бесплатно (fb2, epub, mobi)
7 ноября, 2021«Готическая проза», «черная романтика», литература «сумеречного настроения»… Как только не называют прозу, вовлекающую читателя в… >>>Пушкин А.С. Арап Петра Великого. Скачать бесплатно (fb2, epub, mobi)
13 февраля, 2021«В числе молодых людей, отправленных Петром Великим в чужие края для приобретения сведений, необходимых государству… >>>Пушкин А.С. Гавриилиада. Скачать бесплатно (fb2, epub, mobi)
25 декабря, 2020 Скачать бесплатно fb2 Скачать бесплатно epub Скачать бесплатно mobi. .. >>>
.. >>>Как познать себя?
16 ноября, 2020В жизни людей наступает момент, когда становится интересно: «А кто я такой и зачем я… >>>Гоголь Н.В. Портрет. Скачать бесплатно
14 октября, 2020Впервые повесть напечатана в «Арабесках» в 1835 г. Над «Портретом» Гоголь работал в течение 1833-1834… >>>
Новые митрофанушки на рынке труда
Литература: Рефлексия. Актуальность творчества Фонвизина на примере образа Митрофанушки из комедии «Недоросль».
Деятельностная цель:
- формирование у учащихся деятельностных способностей и способностей к структурированию и систематизации изучаемого предметного содержания экономики и текстов по литературе;
- формирование способностей учащихся к оценке ситуаций как в жизни, так и в произведениях художественной литературы, умений связать понятия из различных школьных предметов для составления целостного мировоззрения.

Содержательная цель:
- построение обобщенных понятий о ценности знаний, получаемых в школе, и выявление закономерностей в поведении человека на рынках товаров и рынке труда;
- развитие речи и умений учащихся формировать и выражать свою позицию на основе полученных знаний;
- совершенствование умения выразительного ролевого и анализирующего чтения.
Тип урока: урок-исследование.
Оборудование урока: текст комедии «Недоросль», интерактивная доска, раздаточный материал.
Структура урока:
- Актуализация знаний.
- Приветствие учителя. Объявление темы урока. – 1 мин.
- Диалог госпожи Простаковой и учителя экономики. 1 мин.
- Постановка задачи, выявление цели урока.
- чтение по ролям (действие III явление VII) 6 мин.
- Слайды-сценки 2 мин
- Слайды (реклама) 1 мин.
- Определение цели урока 1 мин.
- Слайды со списком товаров – определение компетенций покупателя 5 мин.
- Слайды со схемой и анализ раздаточного материала по вакансиям 5 мин.
- чтение по ролям (действие III явление VII) 6 мин.
- Оценочно-рефлексивный этап
- Интерпретация полученных данных 5 мин.
- Вывод по результатам исследовательской работы 5 мин.
- Применение новых знаний в учебной деятельности 5 мин.
- Итоги урока и выставление оценок 3 мин.
- Резерв времени 5 мин.
Ход урока
Учитель экономики: Приветствие.
– Кто такие Митрофанушка, госпожа Простакова, Цыфиркин, Кутейкин, Вральман? (герои комедии Д.И. Фонвизина «Недоросль») Кратко охарактеризуйте их.
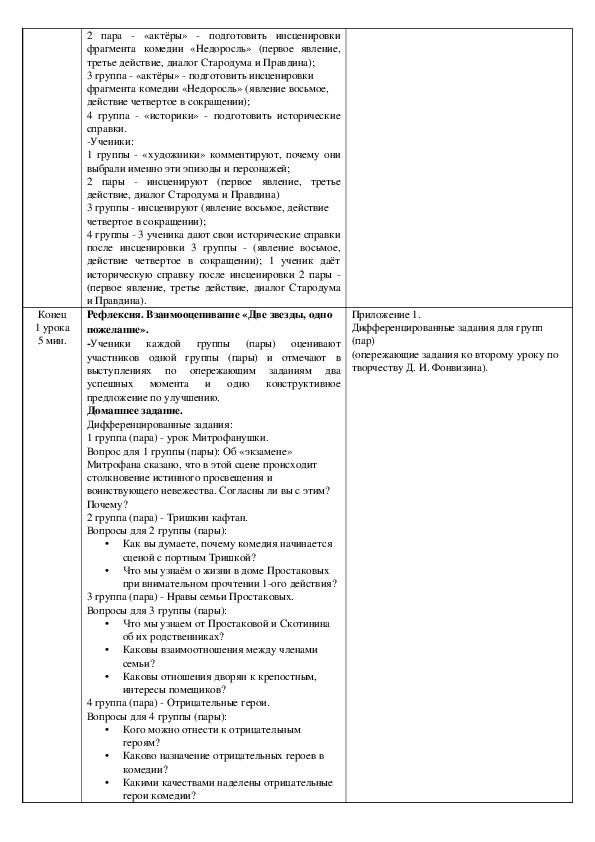
Ответы: Госпожа Простакова – провинциальная помещица, в девичестве Скотинина, властная, бесцеремонная и деспотичная госпожа, грубая и невежественная женщина, но при этом нежная и любящая мать.
Митрофан – сын Простаковой, шестнадцатилетний увалень, невежда, эгоист, лентяй, тунеядец.
Цыфиркин (Пафнутьич) – учитель Митрофана, отставной солдат, трудолюбив, честен. Он пытается научить Митрофана азам математики, получает за свой труд от Простаковой жалкие гроши.
Кутейкин (Сидорыч) – недоучившийся семинарист, жадный и хитрый человек, обучал Митрофана грамматике русского и церковнославянского языков.
Вральман Адам Адамыч – бывший кучер Стародума, невежественный немец, хвастливый и амбиционный, мошенник, обучал Митрофана «по-французски и всем наукам».
– А вот и сама госпожа Простакова. Сударыня, скажите, как ваши родители относились к образованию?
Простакова: Не нынешний был век. Нас ничему не учили. Бывало, добры люди приступят к батюшке, ублажают, ублажают, чтоб хоть братца отдать в школу. К статью ли, покойник-свет и руками и ногами, царство ему небесное! Бывало, изволит закричать: прокляну робенка, который что-нибудь переймет у басурманов, и не будь тот Скотинин, кто чему-нибудь учиться захочет.
Учитель экономики: Вы, однако ж, своего сынка кое-чему обучаете.
Простакова: Да ныне век другой, матушка! Последних крох не жалеем, лишь бы сына всему выучить. Митрофанушка, моя забота, моя отрада, уж четыре года как учится, из-за книги не встает по суткам. Материно мое сердце. Иное жаль, жаль, да подумаешь: зато будет и детина хоть куда. Вить вот уж ему, матушка, шестнадцать лет исполнится около зимнего Николы. Из нашей же фамилии Простаковых, смотри-тка, на боку лёжа, летят себе и чины.
Учитель экономики: Ребята, давайте заглянем на урок к Митрофанушке. (Чтение по ролям действие III явление VII)
Учитель литературы: Как вы думаете, заинтересован ли Митрофан в получении знаний? Какими качествами обладает этот ученик?
Учитель экономики: Можно ли доверить Митрофанушке расчёт стоимости покупок и проверку сдачи в магазине?
Ответ. Конечно, нет.
Учитель литературы: Уже при жизни Д.И. Фонвизина имя «Митрофан» стало нарицательным, обозначавшим лентяя и невежду. Заслужить такое прозвище означало опозориться. Говорят, один из молодых дворян узнал себя в Митрофане и до того был потрясён, что взялся за учёбу: изучил языки, философию, историю, живопись; впоследствии стал художником, президентом Академии художеств. Это был Алексей Николаевич Оленин – один из образованнейших людей своего времени.
Учитель экономики: К сожалению, и сегодня можно встретить некоторых учеников, которые не слишком усердно осваивают науки.
Просмотр слайдов (сценка о русском языке) с комментариями учителя литературы.
Учитель экономики: А теперь давайте попробуем сформулировать цели нашего сегодняшнего урока.
Ответ. Мы попытаемся выяснить, является ли учёба в школе основой будущего успеха в профессии и может ли рассчитывать человек с низким качеством образования на хорошую зарплату.
Учитель экономики: Можно ли доверить митрофанушкам покупку сложных технических товаров: например, видеокамеры, многофункциональной стиральной машины?
Ответ. Они не смогут разобраться в инструкции и правильно использовать приборы.
Учитель экономики: Может ли неграмотный человек стать рациональным потребителем, грамотным покупателем?
Ответ. Нет
Нет
Учитель литературы: В том, что русский язык и математику очень важно в школе добросовестно изучать (знания этих учебных предметов всегда пригодятся в жизни), мы убедились. А как же другие предметы?
Просмотр слайдов (сценка о необходимости знаний по географии) с комментариями учителя
Учитель
Ответ.
Вспомним урок на тему «Рынок труда» (слайд)
– Кто на этом рынке покупатель, а кто продавец?
Какой товар хочет купить покупатель?
А что является предметом купли– продажи на этом рынке? (слайд)
Ответ. Объектом купли-продажи на нем является право на использование рабочей силы, знаний, квалификации и способностей к трудовому процессу.
Учитель экономики: Современным митрофанушкам работодатель будет предлагать среднерыночную зарплату?
Ответ. Нет
– Значит, они попадают в зону безработицы, так как считают себя умными и образованными.
А если они по знакомству устраиваются на престижную работу, какого качества изделия могут произвести? Например, такие рекламные щиты могут создать горе работнички. (слайд)
А теперь давайте вспомним требования, которые предъявляются к соискателям таких профессий, как авиакассир и монтажник. (слайд)
-
Авиакассир/менеджер по продаже авиа и железнодорожных билетов
Опыт работы от 2-х лет, знание систем Galileo, Сирена, СПЖД, Gabriel.
Исполнительность, доброжелательность, грамотная речь
хорошие навыки устного счета. -
Монтажник
Опыт работы на монтаже технологических трубопроводов и оборудования‚ Знание требований техники безопасности.
Итак, кто скажет, какие компетенции, умения требуются в этих профессиях?
Ответ. Грамотная речь, математические навыки, компьютерная грамотность.
– А как себя будут чувствовать новые митрофанушки на рынке труда с теми знаниями, которые имеют?
Ответ. Страна, в которой малообразованное население преобладает, не производит сложных товаров, а как следствие —
- плохие зарплаты
- бедность
- воровство, бандитизм
- и даже революции
Учитель экономики: Снова рассмотрите раздаточный материал и скажите, какой уровень знаний родного языка и математики должен быть у людей самых высокооплачиваемых профессий?
Ответ. Высокий.
Учитель литературы: К соискателям разных профессий предъявляются следующие требования: быть коммуникабельным, культурным в общении, доброжелательным.
А как ведет себя Митрофанушка?
Ответ.
Учитель экономики: Культура поведения – тоже конкурентное преимущество на рынке труда.
Теперь давайте подведём итоги. Как освоение знаний в школе влияет на будущую жизнь человека, на жизнь страны в целом?
Кто хочет высказать свое мнение?
Ответ.
Учитель литературы: Почему комедия «Недоросль», написанная Д.И. Фонвизиным в 1781(!) году, и сегодня актуальна?
Ответ.
– Д.И. Фонвизин надеялся на то, что придёт время, когда молодёжь будет считать позором бездеятельную жизнь: «Дворянин считал бы за первое бесчестие не делать ничего, когда есть ему столько дела, есть люди, которым помогать надо, есть Отечество, которому служить», и в результате Россия станет образованной, независимой, духовно богатой, ибо достоинство нации определяется делами её народа. Комедия «Недоросль» – современно звучащее произведение, помогающее нам вести непримиримый бой с ещё встречающимися митрофанушками, простаковыми, скотиниными, которые тормозят экономическое, духовное, культурное развитие России.
Решение кроссворда.
Проверка кроссворда.
Учитель экономики: Кто перечислит ключевые слова по экономике, которые нам сегодня понадобились, когда мы разбирались с мотивацией к учению?
Ответ.
Выставление оценок.
Домашнее задание по экономике: …
Домашнее задание по литературе: написать эссе «Как я понимаю строки из стихотворения А.С. Пушкина «Послание к цензору»:
Старик превосходный
Невежество казнил
В комедии народной…
Дополнительный материал по литературе:
– Почему отец госпожи Простаковой не хотел, чтобы его дети стали образованными, а сама помещица всё-таки пожелала дать какое-никакое образование своему единственному сыну?
Указом 1714 года Пётр I повелел всем дворянам служить в военной или гражданской службе. Не получившие образования дворяне на службу не принимались, и им запрещалось жениться. «Недорослями» официально называли молодых дворян, ещё не имевших свидетельств об образовании, без которых нельзя было поступить на службу.
«Недорослями» официально называли молодых дворян, ещё не имевших свидетельств об образовании, без которых нельзя было поступить на службу.
– Кому из персонажей комедии «Недоросль» на заявлении написали: «От всякого учения уволить: писано то есть – не мечите бисера перед свиньями, да не попрут его ногами»?
Литература:
- Д.И. Фонвизин «Недоросль» – М.: Просвещение, 2011.
- В.Я. Коровина «Литература 8 класс» – М.: 2011.
- В.С. Автономов «Введение в экономику» – М.: Московский учебник, 2006.
- Сборник заданий «Прикладная Экономика» – Чехов: Достижения молодых, 2006.
Приложения
Афоризмы из комедии Д.И.Фонвизина Недоросль (Сочинение)
«Всякая вина виновата», Тарас Скотинин (действие 1 явление 4)
«Дела не делай, от дела не бегай» (Цыфиркин действие 3 явление 6),
«Дал мне Бог ученичка, боярского сынка» (Цыфиркин действие 3, явление 6),
«виной залетишь за тридевять земель, за тридесято царство» (Правдин, явление 8, действие 4)
«Суженого конём не объедешь» (Скотинин, явление 3, действие 2)
«С великим просвещение можно быть мелкому скареду1» (Стародум, явление 1 действие 3),
«На своё счастье грех пенять» (Скотинин, явление 3, действие 2)
«Век живи, век учись» (Г-жа Простакова, явление 6 действие 2)
«Ученье — вздор» (Скотинин, явление 8, действие 4)
«Где гнев, тут и милость» (Г-жа Простакова, явление 9, действие 4)
«дай мне сроку, хотя на три дня» (Г-жа Простакова, явление 5, действие 5)
«Повинную голову меч не сечёт» (Г-жа Простакова, явление 4, действие 5)
«В большом свете водятся премелкие души» (Стародум, явление 1, действие 3)
«Честнее быть без вины обойдёну, нежели без заслуг пожаловану» (Стародум, явление 1, действие 3)
«Без вины виноват» (Простаков, явление 3, действие 5)
«Без знатных дел знатное состояние ничто» (Стародум, явление 2, действие 4)
«Собака лает, ветер носит» (Цыфиркин, явление 7 действие 3)
«Угнетать рабством себе подобных беззаконно» (Стародум, явление 1, действие 5)
«Сон в руку» (Простаков, явление 4, действие 1)
«Концы в воду» (Скотинин, явление 5, действие 1)
«Видали виды» (Простаков, явление 8, действие 1)
«Белены объелся» (Митрофан, явление 4, действие 2)
«Поминай, как звали» (Митрофан, явление 6, действие 2)
«С его кулаков да за часослов» (Митрофан, явление 6, действие 2)
«Весёлым пирком, да за свадебку» (Скотинин, явление 5, действие 3)
«По добру, по здорову» (Скотинин, явление 4, действие 5)
«Наличные деньги — не наличные достоинства» (Стародум, явление 2, действие 3)
«Ему шабаш из-за доски, а меня рады в толчки» (Цыфиркин, явление 6, действие 3)
Интересно
противопоставлены афоризмы и пословицы
в структуре комедии Д. И. Фонвизина
«Недоросль» (это отметил в своём анализе
речи персонажей литературовед К.Пигарёв).
Речь отрицательных персонажей
(Простаковой, Скотинина) часто неграмотна
и груба, но в ней встречаются народные
выражения и пословицы. А положительные
персонажи со своей культурной, книжной,
построенной на французский манер речью
пословиц почти не употребляют — зато,
скажем, Стародум так и сыплет сентенциями.
И. Фонвизина
«Недоросль» (это отметил в своём анализе
речи персонажей литературовед К.Пигарёв).
Речь отрицательных персонажей
(Простаковой, Скотинина) часто неграмотна
и груба, но в ней встречаются народные
выражения и пословицы. А положительные
персонажи со своей культурной, книжной,
построенной на французский манер речью
пословиц почти не употребляют — зато,
скажем, Стародум так и сыплет сентенциями.
Биоинспирированный посев биоматериалов с использованием трехмерных микротканей вызывает дифференцировку хондрогенных стволовых клеток и образование хрящей в условиях отсутствия факторов роста
Контролируемая высокопроизводительная генерация микроагрегатов стволовых клеток Рис. 1А). Конструкция пластины состояла из массива из 125 000 идеальных кругов диаметром 200 микрометров (дополнительный рис.1А). Круги располагались на расстоянии 100 микрометров друг от друга. Затем была создана негативная копия с использованием полидиметилсилоксана, который служил шаблоном для отливки дисков из 3-процентной агарозы, что позволяет создать микротехнологическую неприлипающую поверхность для культивирования тысяч агрегатов.
 С помощью перфораторов для биопсии были получены микролуночные вставки, подходящие для стандартных культуральных планшетов (дополнительная рис. 1B). 24-луночная микролуночная вставка содержит 2000 отдельных микролунок. Сканирующая электронная микроскопия продемонстрировала высокую точность передачи паттерна процедуры, в результате чего были получены почти идеальные круги со слегка закругленным дном (рис.1Б). Периостальные стволовые клетки человека высевали на вставки микролунки предварительно выбранной высоты в диапазоне от 50 до 250 микрометров. Световой микроскопический анализ показал, что глубина 150 микрометров была оптимальной для образования микроагрегатов и обеспечения возможности их последующего культивирования in vitro (рис. 1С). В частности, микролунки меньшей глубины позволяли формировать микроагрегаты, но не препятствовали миграции новообразованных микротканей через микролунки. Напротив, микроячейки глубиной 150 микрометров и более сохраняли микроагрегаты в течение как минимум 21 дня в культуре (рис.
С помощью перфораторов для биопсии были получены микролуночные вставки, подходящие для стандартных культуральных планшетов (дополнительная рис. 1B). 24-луночная микролуночная вставка содержит 2000 отдельных микролунок. Сканирующая электронная микроскопия продемонстрировала высокую точность передачи паттерна процедуры, в результате чего были получены почти идеальные круги со слегка закругленным дном (рис.1Б). Периостальные стволовые клетки человека высевали на вставки микролунки предварительно выбранной высоты в диапазоне от 50 до 250 микрометров. Световой микроскопический анализ показал, что глубина 150 микрометров была оптимальной для образования микроагрегатов и обеспечения возможности их последующего культивирования in vitro (рис. 1С). В частности, микролунки меньшей глубины позволяли формировать микроагрегаты, но не препятствовали миграции новообразованных микротканей через микролунки. Напротив, микроячейки глубиной 150 микрометров и более сохраняли микроагрегаты в течение как минимум 21 дня в культуре (рис. 1Д). Варьируя плотность засева клеток, мы могли генерировать микроагрегаты различных размеров. В частности, микроагрегаты, содержащие от 50 до 500 клеток, можно воспроизводить, культивировать и собирать для анализа (рис. 1Е). Микроагрегаты, состоящие из большего количества клеток, были успешно получены, но их нельзя было культивировать без дополнительных этапов обработки, поскольку это истощало питательные вещества среды менее чем за 24 часа. Высокая выживаемость клеток в микроагрегатах из 50, 100 и 250 клеток была подтверждена тем, что было обнаружено, что только ~1.5% клеток подверглись апоптозу через семь дней культивирования (рис. 1F, G). Рисунок 1
1Д). Варьируя плотность засева клеток, мы могли генерировать микроагрегаты различных размеров. В частности, микроагрегаты, содержащие от 50 до 500 клеток, можно воспроизводить, культивировать и собирать для анализа (рис. 1Е). Микроагрегаты, состоящие из большего количества клеток, были успешно получены, но их нельзя было культивировать без дополнительных этапов обработки, поскольку это истощало питательные вещества среды менее чем за 24 часа. Высокая выживаемость клеток в микроагрегатах из 50, 100 и 250 клеток была подтверждена тем, что было обнаружено, что только ~1.5% клеток подверглись апоптозу через семь дней культивирования (рис. 1F, G). Рисунок 1 Разработка высокопроизводительной платформы микроагрегации.
( A ) Фотолитографию использовали для создания рисунка микростолбиков на кремниевой пластине, которая действует как мастер-форма. ( B ) СЭМ-микрофотография агарозного слепка эталонной формы продемонстрировала массивы микролунок высокого качества. ( C ) Стволовые клетки надкостницы человека высевали в микролунки глубиной 66 или 150 микрометров ( D ) и оставляли для культивирования минимум на 21 день.( E ) Изменяя плотность посева клеток, можно получить микроагрегаты от 50 до 500 клеток с высокой степенью контроля. Чтобы определить процент апоптотических клеток, микроагрегаты окрашивали с использованием ( F ) мечения концевой дезоксинуклеотидилтрансферазы dUTP, которое было полуколичественно определено ( G ). Масштабные полосы равны 200 мкм.
( C ) Стволовые клетки надкостницы человека высевали в микролунки глубиной 66 или 150 микрометров ( D ) и оставляли для культивирования минимум на 21 день.( E ) Изменяя плотность посева клеток, можно получить микроагрегаты от 50 до 500 клеток с высокой степенью контроля. Чтобы определить процент апоптотических клеток, микроагрегаты окрашивали с использованием ( F ) мечения концевой дезоксинуклеотидилтрансферазы dUTP, которое было полуколичественно определено ( G ). Масштабные полосы равны 200 мкм.
Микроагрегация снижает экспрессию маркеров стволовости
Микроагрегация обеспечивает стволовым клеткам конденсирующее трехмерное микроокружение, напоминающее развивающийся зачаток конечности.Ключевой особенностью хондрогенной детерминации стволовых клеток зачатков конечностей является снижение их маркеров стволовости. Действительно, воздействие TGFB на периостальные стволовые клетки снижало экспрессию CD73 и CD90 на клеточной поверхности в ~3 и ~2 раза соответственно. Интересно, что микроагрегация более сильно снижала поверхностную экспрессию этих биомаркеров, чем TGFB, примерно в 6 и 7 раз соответственно. Более того, сочетание микроагрегации и воздействия TGFB дополнительно снижало экспрессию CD73 и CD90 в 10 и 9 раз соответственно (рис.2А). Этот эффект был, по крайней мере, частично объяснен изменениями уровней мРНК, поскольку как TGFB, так и микроагрегация приводят к более низким уровням экспрессии генов NT5E и THY1 , которые кодируют CD73 и CD90 соответственно (фиг. 2B). Микроагрегация не влияла ни на экспрессию CD105 на клеточной поверхности, ни на уровни экспрессии мРНК его кодирующего гена ENG . Это можно объяснить относительно низким базальным уровнем экспрессии CD105 в использованных периостальных стволовых клетках.
Интересно, что микроагрегация более сильно снижала поверхностную экспрессию этих биомаркеров, чем TGFB, примерно в 6 и 7 раз соответственно. Более того, сочетание микроагрегации и воздействия TGFB дополнительно снижало экспрессию CD73 и CD90 в 10 и 9 раз соответственно (рис.2А). Этот эффект был, по крайней мере, частично объяснен изменениями уровней мРНК, поскольку как TGFB, так и микроагрегация приводят к более низким уровням экспрессии генов NT5E и THY1 , которые кодируют CD73 и CD90 соответственно (фиг. 2B). Микроагрегация не влияла ни на экспрессию CD105 на клеточной поверхности, ни на уровни экспрессии мРНК его кодирующего гена ENG . Это можно объяснить относительно низким базальным уровнем экспрессии CD105 в использованных периостальных стволовых клетках.
Микроагрегация и TGFB снижают экспрессию маркеров стволовости.
( A ) Одиночные или микроагрегированные hPDC, культивированные в течение семи дней в присутствии или в отсутствие 10 нг/мл TGFB, анализировали на CD73, CD90 и CD105 с использованием FACS и ( B ) на экспрессию их соответствующих гены NTE5 , THY1 и ENG с использованием RT-qPCR. Данные представляют собой среднее значение трех доноров, каждое из которых измерено в четырех повторностях ± ± стандартное отклонение.*P < 0,05; **Р < 0,01; ***Р < 0,001.
Данные представляют собой среднее значение трех доноров, каждое из которых измерено в четырех повторностях ± ± стандартное отклонение.*P < 0,05; **Р < 0,01; ***Р < 0,001.
Микроагрегаты обеспечивают индуцированную TGFB ядерную транслокацию SOX9
Высокая плотность клеток и компактная трехмерная природа микроагрегатов потенциально могут замедлять или даже предотвращать диффузию экзогенно добавленных факторов роста в ядро клеточной конструкции. Чтобы исследовать это, мы подвергли микроагрегаты, состоящие из 50, 100 или 250 клеток, воздействию TGFB и сравнили их ядерную транслокацию SOX9 с транслокацией обычных монослоев.С этой целью микроагрегаты были успешно визуализированы сверху вниз с помощью флуоресцентной конфокальной микроскопии и представлены в виде сложенных изображений (рис. 3). Опосредованная SOX9 транскрипция является ключевым событием в хондрогенезе. Микроагрегаты из 50 и 100 клеток продемонстрировали аналогичный процент клеток с SOX9-положительным ядром, который колебался от 30 до 35 процентов. Примечательно, что часть клеток в микроагрегатах окрашивалась заметно интенсивнее по сравнению с их монослойными аналогами.50 клеточных микроагрегатов продемонстрировали SOX9-положительные клетки во всех микротканях. Напротив, 100-клеточные микроагрегаты отдавали небольшое предпочтение более периферической области и меньше — ядру микроткани. Кроме того, микроагрегаты из 250 клеток имели значительно меньшую долю клеток, демонстрирующих ядерную транслокацию SOX9 (~ 17%), которые обычно располагались в более периферических областях. В монослойных культурах без TGFB процент положительных по SOX9 клеток был незначительным.Поразительно, но микроагрегированные культуры без TGFB содержали до 10% SOX9-позитивных клеток. Это явление зависело от размера микроагрегата с более высоким процентом SOX9-положительных клеток, а также с более интенсивно окрашенными клетками в более мелких микроагрегатах.
Примечательно, что часть клеток в микроагрегатах окрашивалась заметно интенсивнее по сравнению с их монослойными аналогами.50 клеточных микроагрегатов продемонстрировали SOX9-положительные клетки во всех микротканях. Напротив, 100-клеточные микроагрегаты отдавали небольшое предпочтение более периферической области и меньше — ядру микроткани. Кроме того, микроагрегаты из 250 клеток имели значительно меньшую долю клеток, демонстрирующих ядерную транслокацию SOX9 (~ 17%), которые обычно располагались в более периферических областях. В монослойных культурах без TGFB процент положительных по SOX9 клеток был незначительным.Поразительно, но микроагрегированные культуры без TGFB содержали до 10% SOX9-позитивных клеток. Это явление зависело от размера микроагрегата с более высоким процентом SOX9-положительных клеток, а также с более интенсивно окрашенными клетками в более мелких микроагрегатах.
Ядерная транслокация SOX9 в одиночных и микроагрегированных hPDC.
hPDC, культивированные в виде отдельных клеток или микроагрегатов в течение семи дней в присутствии или в отсутствие 10 нг/мл TGFB, были иммуногистохимически окрашены на SOX9, микрофотографированы и полуколичественно определены.Масштабные полосы равны 200 мкм. **Р < 0,01; ***P < 0,001
Микроагрегация улучшает хондрогенный транскрипционный профиль
Для определения влияния микроагрегации на хондрогенную дифференцировку уровней мРНК периостальных стволовых клеток количественно определяли после 6 дней культивирования (рис. 4). Совпадая с иммуногистохимией SOX9, добавление TGFB и микроагрегация, в частности с использованием 50 и 100 клеток, повышали уровни мРНК SOX9 , COL2A1 и ACAN .мРНК COL10A1 незначительно увеличивалась при TGFB и более сильно при микроагрегации. Хотя TGFB, безусловно, является наиболее часто используемым фактором роста для индукции хондрогенной дифференцировки, ранее мы сообщали, что in vivo периостальных стволовых клеток мышей превращаются в хондрогенную линию костного морфогенетического белка (BMP) 2 16 . Интересно, что в то время как добавка TGFB снижала экспрессию мРНК BMP2 в отдельных клетках, микроагрегация сильно увеличивала уровни мРНК BMP2 в зависимости от размера микроагрегатов.Чтобы выяснить, имело ли это измеримый последующий эффект, мы измерили уровни мРНК прямого гена-мишени BMP ID1 , которые тесно следовали за экспрессией BMP2. Индуцируемая BMP2 остеохондральная дифференцировка действует через RUNX2 и опосредована через DLX5. Уровни мРНК этих двух генов не претерпели значительных изменений при добавлении TGFB, но значительно увеличились при микроагрегации независимо от их размера. Более того, микроагрегация, но не добавление TGFB, увеличивала уровни мРНК OSX и OCN .Интересно, что уровни экспрессии мРНК COL1 повышались при воздействии TGFB в контрольных одиночных клетках, однако это сводилось на нет при микроагрегации. Кроме того, добавление TGFB сильно снижало экспрессию мРНК ALPL , которая не усиливалась микроагрегацией.
Интересно, что в то время как добавка TGFB снижала экспрессию мРНК BMP2 в отдельных клетках, микроагрегация сильно увеличивала уровни мРНК BMP2 в зависимости от размера микроагрегатов.Чтобы выяснить, имело ли это измеримый последующий эффект, мы измерили уровни мРНК прямого гена-мишени BMP ID1 , которые тесно следовали за экспрессией BMP2. Индуцируемая BMP2 остеохондральная дифференцировка действует через RUNX2 и опосредована через DLX5. Уровни мРНК этих двух генов не претерпели значительных изменений при добавлении TGFB, но значительно увеличились при микроагрегации независимо от их размера. Более того, микроагрегация, но не добавление TGFB, увеличивала уровни мРНК OSX и OCN .Интересно, что уровни экспрессии мРНК COL1 повышались при воздействии TGFB в контрольных одиночных клетках, однако это сводилось на нет при микроагрегации. Кроме того, добавление TGFB сильно снижало экспрессию мРНК ALPL , которая не усиливалась микроагрегацией. Кроме того, чтобы убедиться, что улучшенный профиль экспрессии хондрогенных генов сохраняется с течением времени, отдельные клетки и микроагрегаты из 100 клеток анализировали на экспрессию генов после культивирования в присутствии TGFB в течение 21 дня (фиг.5А). Действительно, по сравнению с одиночными клетками микроагрегация увеличивалась SOX9 (~1,5 раза), ACAN (~2,5 раза) и COL2A1 (~6 раз), уменьшалась COL1 (~2,5 раза). 5 раз) и не изменил уровни экспрессии мРНК COL10A1 . Этот паттерн экспрессии генов указывает на то, что микроагрегация более способствует хондрогенной дифференцировке, чем отдельные клетки. Кроме того, мы определили, что отложение коллагена стало значительно выше в микроагрегатах по сравнению с отдельными клетками, примерно в 2,5 и 6,5 раза выше после 12 и 21 дней культивирования (рис.5Б). Точно так же отложение ГАГ было значительно выше в микроагрегатах, чем в одиночных клетках, примерно в 2 и 3,5 раза после 12 и 21 дней культивирования (рис.
Кроме того, чтобы убедиться, что улучшенный профиль экспрессии хондрогенных генов сохраняется с течением времени, отдельные клетки и микроагрегаты из 100 клеток анализировали на экспрессию генов после культивирования в присутствии TGFB в течение 21 дня (фиг.5А). Действительно, по сравнению с одиночными клетками микроагрегация увеличивалась SOX9 (~1,5 раза), ACAN (~2,5 раза) и COL2A1 (~6 раз), уменьшалась COL1 (~2,5 раза). 5 раз) и не изменил уровни экспрессии мРНК COL10A1 . Этот паттерн экспрессии генов указывает на то, что микроагрегация более способствует хондрогенной дифференцировке, чем отдельные клетки. Кроме того, мы определили, что отложение коллагена стало значительно выше в микроагрегатах по сравнению с отдельными клетками, примерно в 2,5 и 6,5 раза выше после 12 и 21 дней культивирования (рис.5Б). Точно так же отложение ГАГ было значительно выше в микроагрегатах, чем в одиночных клетках, примерно в 2 и 3,5 раза после 12 и 21 дней культивирования (рис. 5С).
5С).
Микроагрегация индуцирует отпечаток экспрессии хондрогенного гена.
hPDC культивировали в виде отдельных клеток или микроагрегатов в присутствии или в отсутствие 10 нг/мл TGFB в течение семи дней и анализировали на экспрессию генов с помощью RT-qPCR. *P < 0,05; **Р < 0,01; ***P < 0,001 по сравнению с одиночными hPDC без TGFB. # P < 0,05; ## P < 0,01; ### P < 0,001 по сравнению с одиночными hPDC с TGFB.
Рисунок 5Микроагрегация приводит к улучшенному отпечатку экспрессии хондрогенного гена через три недели в культуре. hPDC
культивировали как отдельные клетки или микроагрегаты из 100 клеток в течение 21 дня и анализировали на их соответствующую экспрессию генов SOX9 , ACAN , COL2A1 , COL1A1 и
5 COLX
.Кроме того, их ( B ) коллаген и ( C ) гликозаминогликан (GAG) измеряли в различные моменты времени и нормализовали по содержанию их ДНК, что продемонстрировало значительное повышение продукции коллагена и GAG. Данные представлены как среднее ± SD. N = 4.*P < 0,05; **P < 0,01.
Данные представлены как среднее ± SD. N = 4.*P < 0,05; **P < 0,01.Микроагрегация улучшает
in vivo образование хрящаЗатем мы исследовали, улучшает ли также микроагрегация in vivo хондрогенез и последующее образование хряща.С этой целью клетки надкостницы человека, культивированные либо в виде отдельных клеток, либо в виде микроагрегатов в присутствии или в отсутствие TGFB в течение 6 дней, загружали в коллагеновый гидрогель.
Для этих экспериментов были отобраны микроагрегаты из 100 клеток на основании их общей стабильности и хондрогенного поведения. В этот момент ни в одном из состояний не было обнаруживаемых уровней гликозаминогликанов (дополнительная рис. 2). Следовательно, любые обнаруживаемые уровни гликозаминогликанов будут откладываться исключительно во время фазы in vivo .Через неделю после имплантации образцы были извлечены и разрезаны (рис. 6).
Рисунок 6 Формирование хряща в имплантатах, содержащих одиночные или микроагрегированные hPDC, через неделю после имплантации.
После шести дней культивирования in vitro hPDC помещали в гидрогели коллагена с концентрацией 5 мг/мл, имплантировали подкожно на одну неделю и анализировали гистологически для визуализации имплантата и проникновения в перфузированные кровеносные сосуды с использованием трихрома Массона, клеточную морфологию с использованием гематоксилина и эозина и формирование хряща с использованием альцианового синего и COL2 (n = 6).Белые полосы на шкале равны 200 мкм. Черные полосы шкалы равны 1 мм.
Трихром Массона позволил идентифицировать имплантат путем окрашивания гидрогеля коллагена. Кроме того, это окрашивание показало, что все образцы одиночных клеток без TGFB были инвазированы многочисленными мелкими кровеносными сосудами, даже в сердцевине. В меньшей степени это наблюдалось в имплантатах, содержащих микроагрегаты, и не наблюдалось в клетках, подвергшихся воздействию TGFB. Интересно, что за этот короткий промежуток времени микроагрегаты в значительной степени разобрались на отдельные клетки, которые гомогенно мигрировали по всему имплантату. Потенциально это можно объяснить способностью клетки мощно связываться с биоматериалом, что может стимулировать разборку инкапсулированных микроагрегатов. Эти наблюдения были подтверждены с помощью окрашивания гематоксилин-эозином и подчеркивали в целом веретенообразную морфологию клеток во всех имплантатах.
Потенциально это можно объяснить способностью клетки мощно связываться с биоматериалом, что может стимулировать разборку инкапсулированных микроагрегатов. Эти наблюдения были подтверждены с помощью окрашивания гематоксилин-эозином и подчеркивали в целом веретенообразную морфологию клеток во всех имплантатах.
Трихромное окрашивание по Массону выявило многочисленные кровоснабжаемые кровеносные сосуды, которые присутствовали исключительно в одноклеточных имплантатах, не подвергавшихся воздействию TGFB. Кроме того, несколько кровеносных сосудов наблюдались вокруг, но не внутри микроагрегатного имплантата без TGFB.Напротив, в образцах, содержащих TGFB, кровеносных сосудов не наблюдалось. Более того, коллагеновый матрикс, в котором были инкапсулированы одиночные клетки без TGFB, окрашивался заметно менее интенсивно, чем любой другой имплантат. Это потенциально указывает на ремоделирование матрикса, что коррелирует с его интенсивной сосудистой инвазией. Это было подтверждено окрашиванием H&E, которое выявило многочисленные сосуды и жироподобные клетки внутри имплантатов. Напротив, другой образец имел отчетливую морфологию округлых клеток, расположенных в матриксе, фенотипически напоминающих хондроциты.Небольшие трещины в матрице, вероятно, вызваны клеточным изменением исходных веретенообразных клеток в более округлую морфологию.
Напротив, другой образец имел отчетливую морфологию округлых клеток, расположенных в матриксе, фенотипически напоминающих хондроциты.Небольшие трещины в матрице, вероятно, вызваны клеточным изменением исходных веретенообразных клеток в более округлую морфологию.
Как и ожидалось, отдельные клетки без стимуляции TGFB откладывали лишь незначительное количество гликозаминогликанов и COL2, тогда как добавление TGFB усиливало их отложение. Однако, как это типично для стимулированных имплантатов недельной давности, общие уровни депонированных гликозаминогликанов и COL2 оставались низкими. Напротив, микроагрегация периостальных стволовых клеток приводила к значительно более высоким уровням депонированных гликозаминогликанов и COL2 даже в отсутствие TGFB.Через три недели после имплантации отдельные клетки, которые не получали добавку TGFB, не демонстрировали какого-либо положительного окрашивания на гликозаминогликаны и COL2, в то время как имплантированные, которые получали добавку TGFB, продемонстрировали умеренный уровень окрашивания. Несмотря на это, имплантаты, которые изначально были микроагрегированы, окрашивались более интенсивно для COL2. Важно отметить, что микроагрегированные hPDC, которые не подвергались воздействию TGFB, продемонстрировали наиболее интенсивное окрашивание COL2. В совокупности наши данные свидетельствуют о том, что посев биоматериалов с хондрогенной микросредой, обеспечиваемой клеточными микроагрегатами, превосходит обычную загрузку факторами роста с точки зрения формирования хряща (рис.7).
Несмотря на это, имплантаты, которые изначально были микроагрегированы, окрашивались более интенсивно для COL2. Важно отметить, что микроагрегированные hPDC, которые не подвергались воздействию TGFB, продемонстрировали наиболее интенсивное окрашивание COL2. В совокупности наши данные свидетельствуют о том, что посев биоматериалов с хондрогенной микросредой, обеспечиваемой клеточными микроагрегатами, превосходит обычную загрузку факторами роста с точки зрения формирования хряща (рис.7).
Формирование хряща в имплантатах, содержащих одиночные или микроагрегированные hPDC, через три недели после имплантации.
После шести дней культивирования in vitro hPDC помещали в гидрогели коллагена с концентрацией 5 мг/мл, имплантировали подкожно на три недели и анализировали гистологически для визуализации имплантата и проникновения в перфузированные кровеносные сосуды с использованием трихрома Массона, клеточную морфологию с использованием гематоксилина и эозина и формирование хряща с использованием альцианового синего и COL2 (n = 6). Белые полосы на шкале равны 200 мкм. Черные полосы шкалы равны 1 мм.
Белые полосы на шкале равны 200 мкм. Черные полосы шкалы равны 1 мм.
%PDF-1.4
%
3258 0 объект >
эндообъект
внешняя ссылка
3258 368
0000000016 00000 н
0000010246 00000 н
0000010526 00000 н
0000010927 00000 н
0000012458 00000 н
0000012609 00000 н
0000012760 00000 н
0000012911 00000 н
0000013063 00000 н
0000013216 00000 н
0000013368 00000 н
0000013521 00000 н
0000013672 00000 н
0000013825 00000 н
0000013979 00000 н
0000014133 00000 н
0000014287 00000 н
0000014441 00000 н
0000014595 00000 н
0000014749 00000 н
0000014903 00000 н
0000015057 00000 н
0000015210 00000 н
0000015364 00000 н
0000015518 00000 н
0000015672 00000 н
0000015826 00000 н
0000015980 00000 н
0000016133 00000 н
0000016287 00000 н
0000016441 00000 н
0000016595 00000 н
0000016749 00000 н
0000016903 00000 н
0000017057 00000 н
0000017211 00000 н
0000017365 00000 н
0000017519 00000 н
0000017672 00000 н
0000017826 00000 н
0000017979 00000 н
0000018132 00000 н
0000018286 00000 н
0000018439 00000 н
0000018592 00000 н
0000018746 00000 н
0000018900 00000 н
0000019054 00000 н
0000019208 00000 н
0000019362 00000 н
0000019516 00000 н
0000019670 00000 н
0000019824 00000 н
0000019978 00000 н
0000020132 00000 н
0000020286 00000 н
0000020440 00000 н
0000020594 00000 н
0000020748 00000 н
0000020902 00000 н
0000021056 00000 н
0000021209 00000 н
0000021363 00000 н
0000021516 00000 н
0000021670 00000 н
0000021823 00000 н
0000021975 00000 н
0000022128 00000 н
0000022283 00000 н
0000022438 00000 н
0000022592 00000 н
0000022747 00000 н
0000022901 00000 н
0000023056 00000 н
0000023210 00000 н
0000023365 00000 н
0000023519 00000 н
0000023674 00000 н
0000023828 00000 н
0000023983 00000 н
0000024137 00000 н
0000024292 00000 н
0000024447 00000 н
0000024601 00000 н
0000024754 00000 н
0000024908 00000 н
0000025062 00000 н
0000025216 00000 н
0000025371 00000 н
0000025525 00000 н
0000025680 00000 н
0000025834 00000 н
0000025988 00000 н
0000026142 00000 н
0000026297 00000 н
0000026451 00000 н
0000026606 00000 н
0000026760 00000 н
0000026915 00000 н
0000027069 00000 н
0000027224 00000 н
0000027379 00000 н
0000027533 00000 н
0000027688 00000 н
0000027843 00000 н
0000027999 00000 н
0000028154 00000 н
0000028309 00000 н
0000028463 00000 н
0000028618 00000 н
0000028773 00000 н
0000028928 00000 н
0000029083 00000 н
0000029238 00000 н
0000029393 00000 н
0000029548 00000 н
0000029703 00000 н
0000029858 00000 н
0000030012 00000 н
0000030167 00000 н
0000030322 00000 н
0000030477 00000 н
0000030632 00000 н
0000030787 00000 н
0000030942 00000 н
0000031097 00000 н
0000031252 00000 н
0000031407 00000 н
0000031561 00000 н
0000031716 00000 н
0000031871 00000 н
0000032025 00000 н
0000032180 00000 н
0000032335 00000 н
0000032489 00000 н
0000032644 00000 н
0000032799 00000 н
0000032954 00000 н
0000033108 00000 н
0000033262 00000 н
0000033417 00000 н
0000033570 00000 н
0000033725 00000 н
0000033880 00000 н
0000034035 00000 н
0000034190 00000 н
0000034345 00000 н
0000034499 00000 н
0000034654 00000 н
0000034809 00000 н
0000034965 00000 н
0000035120 00000 н
0000035276 00000 н
0000035431 00000 н
0000035587 00000 н
0000035743 00000 н
0000035898 00000 н
0000036053 00000 н
0000036208 00000 н
0000036362 00000 н
0000036517 00000 н
0000036672 00000 н
0000036826 00000 н
0000036981 00000 н
0000037136 00000 н
0000037290 00000 н
0000037445 00000 н
0000037600 00000 н
0000037755 00000 н
0000037911 00000 н
0000038065 00000 н
0000038221 00000 н
0000038376 00000 н
0000039110 00000 н
0000039435 00000 н
0000039871 00000 н
0000040259 00000 н
0000040421 00000 н
0000041117 00000 н
0000041155 00000 н
0000041378 00000 н
0000041613 00000 н
0000041857 00000 н
0000042128 00000 н
0000042206 00000 н
0000042435 00000 н
0000043828 00000 н
0000044674 00000 н
0000045692 00000 н
0000046636 00000 н
0000047594 00000 н
0000047763 00000 н
0000048020 00000 н
0000049077 00000 н
0000049945 00000 н
0000050865 00000 н
0000050920 00000 н
0000050975 00000 н
0000051030 00000 н
0000051085 00000 н
0000051140 00000 н
0000051195 00000 н
0000051250 00000 н
0000051305 00000 н
0000051360 00000 н
0000051415 00000 н
0000051470 00000 н
0000051525 00000 н
0000051580 00000 н
0000051635 00000 н
0000051690 00000 н
0000051745 00000 н
0000051800 00000 н
0000051855 00000 н
0000051910 00000 н
0000051965 00000 н
0000052020 00000 н
0000052075 00000 н
0000052130 00000 н
0000052185 00000 н
0000052240 00000 н
0000052295 00000 н
0000052350 00000 н
0000052405 00000 н
0000052460 00000 н
0000052515 00000 н
0000052570 00000 н
0000052625 00000 н
0000052680 00000 н
0000052735 00000 н
0000052790 00000 н
0000052844 00000 н
0000052899 00000 н
0000052953 00000 н
0000053008 00000 н
0000053062 00000 н
0000053117 00000 н
0000053171 00000 н
0000053226 00000 н
0000053280 00000 н
0000053335 00000 н
0000053389 00000 н
0000053444 00000 н
0000053498 00000 н
0000053553 00000 н
0000053607 00000 н
0000053662 00000 н
0000053716 00000 н
0000053771 00000 н
0000053825 00000 н
0000053880 00000 н
0000053934 00000 н
0000053989 00000 н
0000054043 00000 н
0000054098 00000 н
0000054152 00000 н
0000054207 00000 н
0000054261 00000 н
0000054316 00000 н
0000054370 00000 н
0000054425 00000 н
0000054479 00000 н
0000054534 00000 н
0000054588 00000 н
0000054643 00000 н
0000054697 00000 н
0000054752 00000 н
0000054806 00000 н
0000054861 00000 н
0000054915 00000 н
0000054970 00000 н
0000055024 00000 н
0000055079 00000 н
0000055133 00000 н
0000055188 00000 н
0000055242 00000 н
0000055297 00000 н
0000055351 00000 н
0000055406 00000 н
0000055460 00000 н
0000055515 00000 н
0000055569 00000 н
0000055624 00000 н
0000055678 00000 н
0000055733 00000 н
0000055787 00000 н
0000055842 00000 н
0000055896 00000 н
0000055951 00000 н
0000056005 00000 н
0000056060 00000 н
0000056114 00000 н
0000056169 00000 н
0000056223 00000 н
0000056278 00000 н
0000056332 00000 н
0000056387 00000 н
0000056441 00000 н
0000056496 00000 н
0000056550 00000 н
0000056605 00000 н
0000056659 00000 н
0000056714 00000 н
0000056768 00000 н
0000056823 00000 н
0000056877 00000 н
0000056932 00000 н
0000056986 00000 н
0000057041 00000 н
0000057095 00000 н
0000057150 00000 н
0000057204 00000 н
0000057259 00000 н
0000057313 00000 н
0000057368 00000 н
0000057422 00000 н
0000057477 00000 н
0000057531 00000 н
0000057586 00000 н
0000057640 00000 н
0000057695 00000 н
0000057749 00000 н
0000057804 00000 н
0000057858 00000 н
0000057913 00000 н
0000057967 00000 н
0000058022 00000 н
0000058076 00000 н
0000058131 00000 н
0000058185 00000 н
0000058240 00000 н
0000058294 00000 н
0000058349 00000 н
0000058403 00000 н
0000058458 00000 н
0000058512 00000 н
0000058567 00000 н
0000058621 00000 н
0000058676 00000 н
0000058729 00000 н
0000058784 00000 н
0000058837 00000 н
0000058892 00000 н
0000058947 00000 н
0000059002 00000 н
0000059057 00000 н
0000059112 00000 н
0000059167 00000 н
0000059222 00000 н
0000059277 00000 н
0000059332 00000 н
0000059387 00000 н
0000059442 00000 н
0000059497 00000 н
0000062168 00000 н
0000062223 00000 н
0000062278 00000 н
0000062333 00000 н
0000081232 00000 н
0000081287 00000 н
0000081342 00000 н
0000081397 00000 н
0000081452 00000 н
0000081507 00000 н
0000081562 00000 н
0000081617 00000 н
0000010012 00000 н
0000007815 00000 н
трейлер
]>>
startxref
0
%%EOF
3625 0 объект > поток
xWkPSg~O[kU簋. T a& *rVΈHFv$zѝ.(8B-0Rg;R,{g|>
T a& *rVΈHFv$zѝ.(8B-0Rg;R,{g|>
Абляция семейства генов ретинобластомы дерегулирует контроль G1, вызывая иммортализацию и увеличение оборота клеток в условиях, ограничивающих рост
Аннотация
Супрессор ретинобластомы pRB принадлежит к семейству так называемых карманных белков, которое также включает p107 и p130. Эти
белки могут функционально перекрываться в контроле клеточного цикла и подавлении опухоли.Мы создали изогенный набор эмбриональных
линии стволовых (ES) клеток, несущие одиночные или сложные мутации с потерей функции в семействе генов Rb , включая клеточную линию, полностью лишенную всех трех карманных белков. Ни одна из нокаутирующих комбинаций не пострадала
характеристики роста ЭС клеток; однако одновременное удаление всех трех карманных белков сильно нарушило их
способность к дифференциации. Для сгенерированных генотипов также были получены первичные эмбриональные фибробласты мыши (MEF).Пока
инактивация Rb отдельно не ослабляла реакцию старения MEF, pRB/p107-дефицитных MEF после адаптации к культивированию in vitro,
продолжали расти умеренными темпами. Дополнительная абляция p130 сделала MEF полностью нечувствительными к индуцирующим старение
сигналы и сильно увеличили скорость их пролиферации. Хотя МЭФ с тройным нокаутом сохраняли зависимость от якоря, они
не имели надлежащего контроля G 1 и демонстрировали повышенный оборот клеток в условиях ингибирования роста.
Для сгенерированных генотипов также были получены первичные эмбриональные фибробласты мыши (MEF).Пока
инактивация Rb отдельно не ослабляла реакцию старения MEF, pRB/p107-дефицитных MEF после адаптации к культивированию in vitro,
продолжали расти умеренными темпами. Дополнительная абляция p130 сделала MEF полностью нечувствительными к индуцирующим старение
сигналы и сильно увеличили скорость их пролиферации. Хотя МЭФ с тройным нокаутом сохраняли зависимость от якоря, они
не имели надлежащего контроля G 1 и демонстрировали повышенный оборот клеток в условиях ингибирования роста.
Потеря функции гена-супрессора ретинобластомы, RB , является обычным явлением при развитии многих типов опухолей у человека, включая наследственную ретинобластому и спорадическую легочную опухоль,
рак молочной железы и мочевого пузыря (Harbour et al. , 1988; Lee et al., 1988; Horowitz et al., 1990). Многочисленные биохимические исследования поместили pRB в сердце клеточного механизма, который контролирует переход от G 1 к S фазе клеточного цикла (Weinberg 1995).pRB может существовать в гипер- и гипофосфорилированной формах, причем последняя связывается с классом транскрипционных факторов и ингибирует их.
все вместе известные как факторы E2 (E2F), активность которых необходима для транскрипции генов, необходимых для ДНК.
синтез (Бернардс, 1997; Дайсон, 1998; Невинс, 1998). Фосфорилирование pRB циклинзависимыми киназами (CDK) приводит к диссоциации комплекса pRB/E2F, высвобождая транскрипционную
активность E2F.Активность CDK положительно регулируется гетеродимеризацией циклинами, но также отрицательно регулируется
ингибиторы циклинзависимой киназы (ИЦК). Таким образом, функцию pRB можно описать как переключатель клеточного цикла: циклин D1-стимулируется
киназная активность CDK4 выключает pRB через фосфорилирование; это высвобождает активность E2F и способствует переходу G 1 -S; состоянию pRB-ON- способствует ингибирование активности CDK4 с помощью CKI p16 INK4A , что способствует остановке клеточного цикла.
, 1988; Lee et al., 1988; Horowitz et al., 1990). Многочисленные биохимические исследования поместили pRB в сердце клеточного механизма, который контролирует переход от G 1 к S фазе клеточного цикла (Weinberg 1995).pRB может существовать в гипер- и гипофосфорилированной формах, причем последняя связывается с классом транскрипционных факторов и ингибирует их.
все вместе известные как факторы E2 (E2F), активность которых необходима для транскрипции генов, необходимых для ДНК.
синтез (Бернардс, 1997; Дайсон, 1998; Невинс, 1998). Фосфорилирование pRB циклинзависимыми киназами (CDK) приводит к диссоциации комплекса pRB/E2F, высвобождая транскрипционную
активность E2F.Активность CDK положительно регулируется гетеродимеризацией циклинами, но также отрицательно регулируется
ингибиторы циклинзависимой киназы (ИЦК). Таким образом, функцию pRB можно описать как переключатель клеточного цикла: циклин D1-стимулируется
киназная активность CDK4 выключает pRB через фосфорилирование; это высвобождает активность E2F и способствует переходу G 1 -S; состоянию pRB-ON- способствует ингибирование активности CDK4 с помощью CKI p16 INK4A , что способствует остановке клеточного цикла. Важность этого пути контроля G 1 -S подчеркивается тем фактом, что в большинстве случаев рака человека были обнаружены генетические изменения.
которые благоприятствуют состоянию pRB -OFF-. К ним относятся генетическая инактивация RB при ретинобластоме и многих других видах рака; циклин D1 сверхэкспрессия в карциномах молочной железы, пищевода, головы и шеи; CDK4 амплификация или мутационная активация при меланомах; и аннулирование активности p16 INK4A при меланомах, карциномах поджелудочной железы и мочевого пузыря (Hall and Peters 1996; Sherr 1996).
Важность этого пути контроля G 1 -S подчеркивается тем фактом, что в большинстве случаев рака человека были обнаружены генетические изменения.
которые благоприятствуют состоянию pRB -OFF-. К ним относятся генетическая инактивация RB при ретинобластоме и многих других видах рака; циклин D1 сверхэкспрессия в карциномах молочной железы, пищевода, головы и шеи; CDK4 амплификация или мутационная активация при меланомах; и аннулирование активности p16 INK4A при меланомах, карциномах поджелудочной железы и мочевого пузыря (Hall and Peters 1996; Sherr 1996).
Хотя контрольный путь G 1 -S, описанный выше, обеспечил основу для понимания функции pRB по подавлению опухолей, клеточный цикл
контроль в клетках млекопитающих представляет собой гораздо более сложную схему различных, по крайней мере, частично перекрывающихся и взаимодействующих путей.
которые регулируются механизмами положительной и отрицательной обратной связи и могут модулироваться на многих уровнях. Таким образом, pRB разделяет
его E2F-регулирующая активность с двумя гомологами, p107 и p130 (для обзора см. Mulligan and Jacks 1998; Nevins 1998; Lipinski and Jacks 1999).Три белка имеют обширную структурную гомологию, в первую очередь расположенную в двух областях, А и В, которые вместе образуют
так называемый карманный домен, ответственный за связывание с E2F. Эти домены также образуют сайт связывания для многих вирусных онкобелков.
включая аденовирус E1A, большой Т-антиген обезьяньего вируса 40 и вирус папилломы человека E7, все из которых отменяют взаимодействие
с E2F (ДеКаприо и др., 1988; Уайт и др., 1988; Дайсон и др., 1989).Кроме того, за пределами домена A/B р107 и р130 имеют значительную гомологию, тогда как соответствующие области pRB и даже pRB
спейсерная область, которая разделяет субдомены А и В, кажется уникальной. Подчеркнуто более близкое сходство между p107 и p130.
ряд наблюдений позволяет предположить, что эти два белка могут функционально замещать друг друга.
Таким образом, pRB разделяет
его E2F-регулирующая активность с двумя гомологами, p107 и p130 (для обзора см. Mulligan and Jacks 1998; Nevins 1998; Lipinski and Jacks 1999).Три белка имеют обширную структурную гомологию, в первую очередь расположенную в двух областях, А и В, которые вместе образуют
так называемый карманный домен, ответственный за связывание с E2F. Эти домены также образуют сайт связывания для многих вирусных онкобелков.
включая аденовирус E1A, большой Т-антиген обезьяньего вируса 40 и вирус папилломы человека E7, все из которых отменяют взаимодействие
с E2F (ДеКаприо и др., 1988; Уайт и др., 1988; Дайсон и др., 1989).Кроме того, за пределами домена A/B р107 и р130 имеют значительную гомологию, тогда как соответствующие области pRB и даже pRB
спейсерная область, которая разделяет субдомены А и В, кажется уникальной. Подчеркнуто более близкое сходство между p107 и p130.
ряд наблюдений позволяет предположить, что эти два белка могут функционально замещать друг друга. р107 и р130 почти
взаимодействуют исключительно с E2F4,5, хотя и на последовательных стадиях клеточного цикла: Комплексы p130 и E2F4,5
преобладают в покоящихся или дифференцированных клетках; Комплексы p107/E2F4,5 обнаруживаются только в циклирующих клетках.Напротив, pRb
имеет самое сильное сродство к E2F1,2,3, хотя он также может связывать E2F4 (для обзора см. Dyson 1998). Вместо того, чтобы просто изолировать E2F, комплексы карманный белок/E2F, по-видимому, активно участвуют в репрессии транскрипции.
(Чжан и др., 1999). В то время как не было обнаружено различий в экспрессии генов-мишеней E2F при удалении только p107 или p130, комбинированная потеря в p107 -/- p130 -/- первичные эмбриональные фибробласты мыши (MEF) привели к дерепрессии ряда генов, включая B-myb, Cdc2, E2f1, Ts, Rrm2 и Cyclin A2 .Экспрессия этих генов была нормальной у Rb -/- MEF, но в этих клетках Cyclin E и p107 были немного активизированы (Herrera et al.
р107 и р130 почти
взаимодействуют исключительно с E2F4,5, хотя и на последовательных стадиях клеточного цикла: Комплексы p130 и E2F4,5
преобладают в покоящихся или дифференцированных клетках; Комплексы p107/E2F4,5 обнаруживаются только в циклирующих клетках.Напротив, pRb
имеет самое сильное сродство к E2F1,2,3, хотя он также может связывать E2F4 (для обзора см. Dyson 1998). Вместо того, чтобы просто изолировать E2F, комплексы карманный белок/E2F, по-видимому, активно участвуют в репрессии транскрипции.
(Чжан и др., 1999). В то время как не было обнаружено различий в экспрессии генов-мишеней E2F при удалении только p107 или p130, комбинированная потеря в p107 -/- p130 -/- первичные эмбриональные фибробласты мыши (MEF) привели к дерепрессии ряда генов, включая B-myb, Cdc2, E2f1, Ts, Rrm2 и Cyclin A2 .Экспрессия этих генов была нормальной у Rb -/- MEF, но в этих клетках Cyclin E и p107 были немного активизированы (Herrera et al. 1996; Hurford et al. 1997). Таким образом, р107 и р130 оказались избыточными в регуляции специфического набора E2F-чувствительных генов, которые не подвержены влиянию.
pRb/E2F-опосредованная репрессия. Однако сверхэкспрессия каждого из карманных белков in vitro может вызвать остановку клеточного цикла.
активность, которая может быть снижена за счет фосфорилирования с помощью cyclinD/CDK4,6.Полная инактивация pRB также требует циклина E/CDK2,
что добавляет дополнительный уровень регуляции пути pRB. Таким образом, три карманных белка могут быть в некоторой степени избыточными.
в блокировании прохождения клеток через клеточный цикл. В покоящихся Т-лимфоцитах основным карманным белком/комплексом E2F является
стр. 130/E2F4. Однако в отсутствие p130 E2F4 в основном находился в комплексе с p107 или, когда этот белок также отсутствовал,
к pRb (Mulligan et al.1998). По-видимому, в этой системе карманные белки могут компенсировать друг друга в регуляции активности E2F4, что объясняет
за их совместную деятельность по блокированию прогрессирования клеточного цикла.
1996; Hurford et al. 1997). Таким образом, р107 и р130 оказались избыточными в регуляции специфического набора E2F-чувствительных генов, которые не подвержены влиянию.
pRb/E2F-опосредованная репрессия. Однако сверхэкспрессия каждого из карманных белков in vitro может вызвать остановку клеточного цикла.
активность, которая может быть снижена за счет фосфорилирования с помощью cyclinD/CDK4,6.Полная инактивация pRB также требует циклина E/CDK2,
что добавляет дополнительный уровень регуляции пути pRB. Таким образом, три карманных белка могут быть в некоторой степени избыточными.
в блокировании прохождения клеток через клеточный цикл. В покоящихся Т-лимфоцитах основным карманным белком/комплексом E2F является
стр. 130/E2F4. Однако в отсутствие p130 E2F4 в основном находился в комплексе с p107 или, когда этот белок также отсутствовал,
к pRb (Mulligan et al.1998). По-видимому, в этой системе карманные белки могут компенсировать друг друга в регуляции активности E2F4, что объясняет
за их совместную деятельность по блокированию прогрессирования клеточного цикла.
Специфические и избыточные функции карманных белков проявлялись также у мышей с одиночными или составными нарушениями в
семейство генов Rb (Mulligan and Jacks 1998). Полная инактивация Rb оказалась несовместимой с нормальным развитием (Clarke et al.1992 год; Джекс и др. 1992 год; Ли и др. 1992). руб -/- эмбрионов, погибших в середине беременности, с обширным апоптозом в центральной нервной системе и хрусталике глаза и дефектами
в эритропоэзе и миогенезе. Эти дефекты могут быть, по крайней мере частично, связаны с повышенной активностью свободного E2F1, индуцирующей
р53-зависимый апоптоз (Morgenbesser et al., 1994; Tsai et al., 1998). Напротив, генетическая абляция ни p107 , ни p130 не выявила явного фенотипа (Cobrinik et al.1996; Ли и др. 1996). Однако комбинация мутаций потери функции p107 и p130 вызывала чрезмерную пролиферацию хондроцитов, приводящую к порокам развития длинных костей и ребер. Проблемы с дыханием, связанные с этим пороком, привели к быстрой неонатальной смерти. Примечательно, что наблюдался драматический фенотип.
когда мутации с потерей функции либо в p107 , либо в p130 были скрещены с фоном BALB/c (LeCouter et al.1998а,б): дефицит р107 вызывал нарушение роста новорожденных, тогда как эмбриональные фибробласты демонстрировали двукратное снижение удвоения
время; Эмбрионы с дефицитом p130 погибали в середине беременности и обнаруживали выраженный апоптоз в центральной нервной системе. Мыши BALB/c были
обнаружено, что он несет точечную мутацию в гене p16 Ink4a (Zhang et al. 1998). Эти наблюдения предполагают, что p107 и p130 могут функционально замещать друг друга, но только в присутствии
путь, который требует p16 дикого типа и, скорее всего, включает pRB.И наоборот, p107, по-видимому, ослаблял последствия
дефицита pRB в развитии, поскольку эмбрионы с дефицитом p107/pRB показали меньшую продолжительность жизни по сравнению с эмбрионами, лишенными
только pRB (Lee et al.
Проблемы с дыханием, связанные с этим пороком, привели к быстрой неонатальной смерти. Примечательно, что наблюдался драматический фенотип.
когда мутации с потерей функции либо в p107 , либо в p130 были скрещены с фоном BALB/c (LeCouter et al.1998а,б): дефицит р107 вызывал нарушение роста новорожденных, тогда как эмбриональные фибробласты демонстрировали двукратное снижение удвоения
время; Эмбрионы с дефицитом p130 погибали в середине беременности и обнаруживали выраженный апоптоз в центральной нервной системе. Мыши BALB/c были
обнаружено, что он несет точечную мутацию в гене p16 Ink4a (Zhang et al. 1998). Эти наблюдения предполагают, что p107 и p130 могут функционально замещать друг друга, но только в присутствии
путь, который требует p16 дикого типа и, скорее всего, включает pRB.И наоборот, p107, по-видимому, ослаблял последствия
дефицита pRB в развитии, поскольку эмбрионы с дефицитом p107/pRB показали меньшую продолжительность жизни по сравнению с эмбрионами, лишенными
только pRB (Lee et al. 1996). Активность p107, по-видимому, также модулирует супрессорную функцию pRB. В отличие от человека, у которого гемизиготность
для гена-супрессора ретинобластомы RB сильно предрасполагает к ретинобластоме, Rb +/− у мышей развились опухоли, происходящие из промежуточной доли гипофиза, в то время как сетчатка оставалась неизменной
(Ху и др.1994 год; Робанус-Маандаг и др. 1994). Не было обнаружено связи предрасположенности к онкогенезу с дефицитом p107 или p130 (Cobrinik et al., 1996; Lee et al., 1996). Однако одновременная инактивация Rb и p107 у химерных мышей приводила к развитию ретинобластомы, что указывает на то, что p107 может действовать как супрессор опухоли в контексте
дефицита pRb (Robanus-Maandag et al., 1998).
1996). Активность p107, по-видимому, также модулирует супрессорную функцию pRB. В отличие от человека, у которого гемизиготность
для гена-супрессора ретинобластомы RB сильно предрасполагает к ретинобластоме, Rb +/− у мышей развились опухоли, происходящие из промежуточной доли гипофиза, в то время как сетчатка оставалась неизменной
(Ху и др.1994 год; Робанус-Маандаг и др. 1994). Не было обнаружено связи предрасположенности к онкогенезу с дефицитом p107 или p130 (Cobrinik et al., 1996; Lee et al., 1996). Однако одновременная инактивация Rb и p107 у химерных мышей приводила к развитию ретинобластомы, что указывает на то, что p107 может действовать как супрессор опухоли в контексте
дефицита pRb (Robanus-Maandag et al., 1998).
Первичные эмбриональные фибробласты мыши представляют собой довольно четко определенный тип клеток, который широко используется для выявления последствий
абляции генов для контроля клеточного цикла. Хотя дефицит pRB или комбинированный дефицит p107/p130 вызывали небольшое ускорение
вступления в S-фазу при рестимуляции клеток, лишенных сыворотки, общие характеристики роста при одиночном или двойном нокауте
MEF не отличались от MEF дикого типа (Cobrinik et al., 1996; Herrera et al., 1996). В частности, испытанные до сих пор генотипы с одинарным и двойным нокаутом не влияли на реакцию остановки роста клеток.
к ряду ингибирующих рост условий, таких как депривация сыворотки, контактное ингибирование и длительное пассирование.Это снова
предполагает функциональную избыточность в семействе карманных белков. Чтобы проверить это напрямую, мы сгенерировали MEF, которые
полностью лишен всех трех карманных белков. Наши результаты идентифицируют карманные белки как критические медиаторы старения.
и реакции клеток на различные ограничивающие рост условия.
Хотя дефицит pRB или комбинированный дефицит p107/p130 вызывали небольшое ускорение
вступления в S-фазу при рестимуляции клеток, лишенных сыворотки, общие характеристики роста при одиночном или двойном нокауте
MEF не отличались от MEF дикого типа (Cobrinik et al., 1996; Herrera et al., 1996). В частности, испытанные до сих пор генотипы с одинарным и двойным нокаутом не влияли на реакцию остановки роста клеток.
к ряду ингибирующих рост условий, таких как депривация сыворотки, контактное ингибирование и длительное пассирование.Это снова
предполагает функциональную избыточность в семействе карманных белков. Чтобы проверить это напрямую, мы сгенерировали MEF, которые
полностью лишен всех трех карманных белков. Наши результаты идентифицируют карманные белки как критические медиаторы старения.
и реакции клеток на различные ограничивающие рост условия.
Результаты
Инактивация Rb, p107 и p130 в ES клетках
Изогенный набор дикого типа ( wt ), руб. −/− , руб. −/− p107 −/− , и Rb −/− p107 −/− p130 -/- линии эмбриональных стволовых (ES) клеток были получены последовательными раундами гомологичной рекомбинации.Поколение руб −/− и руб. −/− p107 -/- Линии ЭС клеток были описаны ранее (Te Riele et al. 1992;Robanus-Maandag et al. 1998). Чтобы инактивировать p130 , мы создали направленный вектор 129p130-pur (рис. 1А). руб. −/− p107 −/− p130 Клетки -/- были получены двумя способами: Сначала p130 инактивировали в Rb -/- p107 −/− Клетки ES и Rb −/− p107 −/− p130 Клетки +/- культивировали при высоких концентрациях пуромицина для отбора на инактивацию оставшегося p130 аллеля дикого типа; во-вторых, оба аллеля p130 были инактивированы в p107 -/- ES клетки, а затем Rb инактивировали с использованием векторов-мишеней 129Rb-hyg и 129Rb-his.
−/− , руб. −/− p107 −/− , и Rb −/− p107 −/− p130 -/- линии эмбриональных стволовых (ES) клеток были получены последовательными раундами гомологичной рекомбинации.Поколение руб −/− и руб. −/− p107 -/- Линии ЭС клеток были описаны ранее (Te Riele et al. 1992;Robanus-Maandag et al. 1998). Чтобы инактивировать p130 , мы создали направленный вектор 129p130-pur (рис. 1А). руб. −/− p107 −/− p130 Клетки -/- были получены двумя способами: Сначала p130 инактивировали в Rb -/- p107 −/− Клетки ES и Rb −/− p107 −/− p130 Клетки +/- культивировали при высоких концентрациях пуромицина для отбора на инактивацию оставшегося p130 аллеля дикого типа; во-вторых, оба аллеля p130 были инактивированы в p107 -/- ES клетки, а затем Rb инактивировали с использованием векторов-мишеней 129Rb-hyg и 129Rb-his. На рисунке 1B показано, что белок p130 не обнаруживался в ткани головного мозга, полученной из p130 . -/- мышь с таким же нарушением в p130 , что указывает на то, что p130 Аллель − можно считать нулевым аллелем. Таким образом, руб. руб. −/− p107 −/− p130 Клетки -/- полностью лишены карманных белков и в дальнейшем будут обозначаться как клетки с тройным нокаутом ( TKO ).Каждый генотип был получен в двух экземплярах независимых раундов гена, ориентированных, начиная от клеток дикого типа. Никто
генерируемые генотипы в ЭС клетках приводят к очевидному фенотипу, указывая, что карманные белка PRB, P107, и
P130 необязательны для нормального роста ЭС клеток.
На рисунке 1B показано, что белок p130 не обнаруживался в ткани головного мозга, полученной из p130 . -/- мышь с таким же нарушением в p130 , что указывает на то, что p130 Аллель − можно считать нулевым аллелем. Таким образом, руб. руб. −/− p107 −/− p130 Клетки -/- полностью лишены карманных белков и в дальнейшем будут обозначаться как клетки с тройным нокаутом ( TKO ).Каждый генотип был получен в двух экземплярах независимых раундов гена, ориентированных, начиная от клеток дикого типа. Никто
генерируемые генотипы в ЭС клетках приводят к очевидному фенотипу, указывая, что карманные белка PRB, P107, и
P130 необязательны для нормального роста ЭС клеток.

Генерация руб — / — p107 — / — p130 — / — ES-клетки и MEFs.( A ) Рестрикционная карта аллеля p130 дикого типа вокруг кодона 405 в сайте Ava I и конструкцией, нацеленной на ДНК 129 p130 -pur. Зонды 1 и 2 обнаруживают модификации по адресу p130 . ( B ) Вестерн-блоттинг p130 в лизатах, приготовленных из p130 +/+ и стр.130 −/− мозговая ткань. ( C ) ПЦР-анализ Rb, p107 и p130 ДНК, выделенной из пассажа 1 wt , Rb −/− , руб. −/− p107 −/− и TKO MEF.ПЦР-продукты, вытекающие из дикого типа и нокаут-аллелей каждого гена указаны открытыми и сплошными стрелками, соответственно. ( D ) Вестерн-блот-анализ PRB, р107, р130, и циклин Е в лизатах, приготовленных из указанного MEFs. Cdk4 служил погрузки
контроль.
( D ) Вестерн-блот-анализ PRB, р107, р130, и циклин Е в лизатах, приготовленных из указанного MEFs. Cdk4 служил погрузки
контроль.
Limited дифференцировка мощность
руб — / — p107 — / — p130 — / — ES-клетки в teratocarcinomas Для того, чтобы изучить вопрос о том карманные белки становятся существенными при дифференциации клеток ES, мы вводили мас , Rb — / — , Rb — / — p107 −/− , и Rb −/− p107 −/− p130 — / — ЭС клетки подкожно голым мышам. В этой среде ЭС клетки образуют опухоли, подобные тератокарциноме, в которых
могут быть идентифицированы типы дифференцированных клеток, включая нейрональные и мышечные клетки. Через три недели после инъекции тератокарциномы
были удалены, и гистологическое исследование. Опухоли производные от wt , Rb −/− , и руб. −/− p107 -/- ES-клетки были очень похожи в отношении их сильно гетерогенного внешнего вида с обширной дифференцировкой нейронов
(Инжир.2А) и поперечно-полосатые мышцы (рис. 2Б). Обширная дифференцировка нейронов была подтверждена окрашиванием нейрон-специфическими антителами против глиальных фибриллярных клеток.
кислый белок (GFAP) и энолаза, специфичная к нейронам (NSE; не показана). Напротив, клеток TKO ES образовали довольно гомогенную опухолевую массу без какой-либо дифференцировки мышечных клеток (рис.
В этой среде ЭС клетки образуют опухоли, подобные тератокарциноме, в которых
могут быть идентифицированы типы дифференцированных клеток, включая нейрональные и мышечные клетки. Через три недели после инъекции тератокарциномы
были удалены, и гистологическое исследование. Опухоли производные от wt , Rb −/− , и руб. −/− p107 -/- ES-клетки были очень похожи в отношении их сильно гетерогенного внешнего вида с обширной дифференцировкой нейронов
(Инжир.2А) и поперечно-полосатые мышцы (рис. 2Б). Обширная дифференцировка нейронов была подтверждена окрашиванием нейрон-специфическими антителами против глиальных фибриллярных клеток.
кислый белок (GFAP) и энолаза, специфичная к нейронам (NSE; не показана). Напротив, клеток TKO ES образовали довольно гомогенную опухолевую массу без какой-либо дифференцировки мышечных клеток (рис. 2C). Розетоподобные структуры и положительное окрашивание антителом p75 LNGFR (Zorick and Lemke 1996) показали, что опухоли TKO преимущественно состоят из нейробластоподобных клеток (рис.2D; не показано). Кроме того, иммуногистохимия с использованием антител, направленных против маркера пролиферации Ki-67, показала, что
доля пролиферирующих клеток в опухолях ТКО была в 15 раз выше, чем в тератокарциномах других генотипов. Эти данные показывают, что в этой системе абляция
из семейства генов Rb отменяет дифференцировку нейронов и мышц.
2C). Розетоподобные структуры и положительное окрашивание антителом p75 LNGFR (Zorick and Lemke 1996) показали, что опухоли TKO преимущественно состоят из нейробластоподобных клеток (рис.2D; не показано). Кроме того, иммуногистохимия с использованием антител, направленных против маркера пролиферации Ki-67, показала, что
доля пролиферирующих клеток в опухолях ТКО была в 15 раз выше, чем в тератокарциномах других генотипов. Эти данные показывают, что в этой системе абляция
из семейства генов Rb отменяет дифференцировку нейронов и мышц.
Ограниченная дифференциация в ТКО тератокарцином. Гистологические срезы тератокарцином окрашивали гематоксилин-эозином. ( A,B ) Расширенная дифференцировка нейронов (N) и мышц (M) в тератокарциномах, полученных с помощью wt ES клеток. Аналогичные результаты были получены с руб. −/− и руб. −/− p107 −/− ES-клетки.( C,D ) Плотноклеточные TKO тератокарциномы лишены мышечной дифференцировки и состоят из примитивных, относительно недифференцированных нейрональных клеток (PN) с
розеткообразные структуры (R). Увеличение ( A,C ) объектива 10×, ( B,D ) объектива 20×.
Аналогичные результаты были получены с руб. −/− и руб. −/− p107 −/− ES-клетки.( C,D ) Плотноклеточные TKO тератокарциномы лишены мышечной дифференцировки и состоят из примитивных, относительно недифференцированных нейрональных клеток (PN) с
розеткообразные структуры (R). Увеличение ( A,C ) объектива 10×, ( B,D ) объектива 20×.
MEF с дефицитом белка кармана, полученные из химерных эмбрионов
Первичные MEF сыграли важную роль в изучении последствий удаления генов для контроля клеточного цикла.Мы получили
изогенный набор MEF от химерных эмбрионов, генерируемых инъекциями бластоцисты мутантных ES клеток. В этих опытах
мы использовали ЭС клетки, которые также содержали ген устойчивости к неомицину под контролем промотора Rosa26 (Friedrich and Soriano 1991; J.-H. Dannenberg и H. te Riele, в процессе подготовки). Кратковременное культивирование в присутствии G418 обеспечило единый метод получения
Препараты MEF, полученные исключительно из ES-клеток.Это было подтверждено путем генотипирования этих клеток с помощью ПЦР на присутствие
аллелей дикого типа и нокаутных аллелей Rb, p107 и p130 (рис. 1C). Присутствие или отсутствие карманных белков было дополнительно подтверждено анализом вестерн-блоттинга (рис. 1D). Нам удалось получить культуры MEF со следующими генотипами: wt, Rb . −/− , руб. −/− p107 −/− , и несколько неожиданно Rb −/− p107 −/− p130 −/− .
В этих опытах
мы использовали ЭС клетки, которые также содержали ген устойчивости к неомицину под контролем промотора Rosa26 (Friedrich and Soriano 1991; J.-H. Dannenberg и H. te Riele, в процессе подготовки). Кратковременное культивирование в присутствии G418 обеспечило единый метод получения
Препараты MEF, полученные исключительно из ES-клеток.Это было подтверждено путем генотипирования этих клеток с помощью ПЦР на присутствие
аллелей дикого типа и нокаутных аллелей Rb, p107 и p130 (рис. 1C). Присутствие или отсутствие карманных белков было дополнительно подтверждено анализом вестерн-блоттинга (рис. 1D). Нам удалось получить культуры MEF со следующими генотипами: wt, Rb . −/− , руб. −/− p107 −/− , и несколько неожиданно Rb −/− p107 −/− p130 −/− .
Карманные белки необходимы для G
1 контроль Пролиферирующие культуры MEF раннего пассажа метили аналогом тимидина BrdU и анализировали с помощью FACS. На фигуре 3А показано, что одновременная инактивация Rb, p107 и p130 сильно снижает процент клеток в G 1 .Это уменьшение фракции G 1 сопровождалось увеличением популяции S-фазы, при этом культур TKO MEF имели до 65% клеток S-фазы. В руб -/- MEFs, наблюдалось лишь незначительное уменьшение популяции G 1 , тогда как нарушение как Rb , так и p107 давало промежуточный фенотип. Вместе с сильно уменьшенным временем удвоения популяции клеток TKO (см.4), эти наблюдения предполагают, что последовательная абляция pRb, p107 и p130 уменьшает продолжительность фазы G 1 клеточного цикла.
Дерегулированный G 1 — контроль в ТКО МЭФ. ( A ) Проценты G 1 , S и G 2 /M фаз клеточного цикла в пролиферирующих MEF дикого типа ( wt ) и дефицитных по карманному белку MEF.Результаты указаны как средние значения трех независимых экспериментов. Планки погрешностей указывают стандартные отклонения среднего. ( B ) Гистограмма FSC-H, показывающая анализ размера клеток MEF с дефицитом Rb, p107 и p130 по сравнению с аналогами wt . Прямой разброс, FSC-H, указывает на диаметр клеток, а сдвиг влево относительно MEF wt представляет более мелкие клетки.
Рисунок 4.
Характеристики роста MEF с дефицитом карманного белка. ( А ) Кривые роста вес , руб −/− , руб. −/− p107 -/- и ТКО МЭФ в проходах 3 (квадраты), 5 (круги) и 7 (треугольники). На рисунке показан представитель трех независимых экспериментов, каждый исполняется в трех экземплярах.( B ) Пролиферация клеток по протоколу на основе 3T9 указанных MEF для 20 пассажей. С интервалом 3,5 дня общее количество клеток трех независимых культур каждого генотипа определяли перед повторным разведением клеток до 3 × 10 5 на шестилуночный планшет (вход). Эксперимент проводился дважды. Столбики погрешностей указывают стандартные отклонения среднего значения.
G 1 укорочение ранее коррелировало с уменьшением размера клеток в некоторых других экспериментальных условиях, таких как сверхэкспрессия
циклина Е (Ohtsubo and Roberts 1993) и сопутствующей инактивации Rb и p21 (Brugarolas et al. 1998). Поэтому мы определили размер MEF дикого типа и мутантов. Пролиферирующие культуры раннего пассажа фиксировали, окрашивали
с пропидиумиодидом (PI) и анализировали с помощью FACS (фиг. 3B). Относительные размеры клеток тесно связаны с долей клеток в G 1 : TKO
1998). Поэтому мы определили размер MEF дикого типа и мутантов. Пролиферирующие культуры раннего пассажа фиксировали, окрашивали
с пропидиумиодидом (PI) и анализировали с помощью FACS (фиг. 3B). Относительные размеры клеток тесно связаны с долей клеток в G 1 : TKO
Интересно отметить, что в клетках, сверхэкспрессирующих циклин-Е или дефицитных по Rb/p21 , укорочение G 1 коррелирует с повышенными уровнями циклина Е или повышенной активностью киназы CDK2. Однако тогда как руб. руб. −/− и руб. −/− p107 -/- MEF показали повышенный уровень циклина E, TKO MEF экспрессировали циклин E на уровнях дикого типа (рис. 1Д). Таким образом, укорочение G 1 не обязательно сопровождается повышенным уровнем циклина Е.
1Д). Таким образом, укорочение G 1 не обязательно сопровождается повышенным уровнем циклина Е.
MEF дикого типа имеют ограниченную способность к росту при длительном пассировании: скорость их роста постепенно снижается до тех пор, пока они, наконец, не
полная остановка с увеличенной, уплощенной морфологией и содержанием 2n ДНК, состояние, которое называется репликативным
старение (Todaro and Green 1963; Campisi 1997).Участие семейства генов Rb в старении оценивали двумя способами. Сначала определили темпы роста мас., руб. −/− , руб. −/− p107 −/− , и ТКО МЭФ при увеличении проходов. На рис. 4А показано, что темпы роста вес. и руб. -/- MEF постепенно снижались при длительном пассировании, и эти клетки начали стареть примерно к седьмому пассажу. руб −/− стр.107 −/− МЭФ показали пятикратное снижение скорости роста; однако они не полностью прекратили пролиферацию на седьмом пассаже (рис.
4А) и явно не приобрели морфологию, подобную старению (не показано). ТКО МЭФ сохраняли наибольшую пролиферативную способность на поздних пассажах, скорость их роста снижалась менее чем в 2 раза.
Во-вторых, долговременную пролиферативную способность MEF оценивали в соответствии с протоколом на основе 3T9 (рис.4Б). Дикий тип, руб. −/− , и руб. −/− p107 −/− МЭФ показали снижение скорости пролиферации до пассажа 11.
На рис. 4А показано, что темпы роста вес. и руб. -/- MEF постепенно снижались при длительном пассировании, и эти клетки начали стареть примерно к седьмому пассажу. руб −/− стр.107 −/− МЭФ показали пятикратное снижение скорости роста; однако они не полностью прекратили пролиферацию на седьмом пассаже (рис.
4А) и явно не приобрели морфологию, подобную старению (не показано). ТКО МЭФ сохраняли наибольшую пролиферативную способность на поздних пассажах, скорость их роста снижалась менее чем в 2 раза.
Во-вторых, долговременную пролиферативную способность MEF оценивали в соответствии с протоколом на основе 3T9 (рис.4Б). Дикий тип, руб. −/− , и руб. −/− p107 −/− МЭФ показали снижение скорости пролиферации до пассажа 11. В то время как Rb Клетки -/- полностью прекратили рост в этом эксперименте, клетки wt в конечном итоге избежали старения в результате генетического изменения (см. также ниже). руб. −/− p107 -/- МЭФ, хотя первоначально подвергались ингибированию роста, не вступали в старение, а устанавливали постоянную пролиферацию
скорость удвоения популяции от одного до двух раз в 3 дня.Кроме того, клеток TKO первоначально показали некоторое снижение скорости роста. Однако это снижение было незначительным, и клетки продолжали пролиферировать.
со скоростью, которая была примерно в три раза выше, чем у двойных нокаутов. Эти результаты демонстрируют, что инактивация Rb сама по себе не ослабляет реакцию MEF на старение; сопутствующая инактивация Rb и p107 , хотя и не устраняет полностью ингибирование роста при культивировании in vitro, придает постоянную способность к пролиферации,
хотя и с умеренной скоростью, в то время как удаление всех членов семейства Rb -генов делает клетки практически нечувствительными к ингибированию роста при культивировании in vitro и сильно увеличивает
скорость их размножения.
В то время как Rb Клетки -/- полностью прекратили рост в этом эксперименте, клетки wt в конечном итоге избежали старения в результате генетического изменения (см. также ниже). руб. −/− p107 -/- МЭФ, хотя первоначально подвергались ингибированию роста, не вступали в старение, а устанавливали постоянную пролиферацию
скорость удвоения популяции от одного до двух раз в 3 дня.Кроме того, клеток TKO первоначально показали некоторое снижение скорости роста. Однако это снижение было незначительным, и клетки продолжали пролиферировать.
со скоростью, которая была примерно в три раза выше, чем у двойных нокаутов. Эти результаты демонстрируют, что инактивация Rb сама по себе не ослабляет реакцию MEF на старение; сопутствующая инактивация Rb и p107 , хотя и не устраняет полностью ингибирование роста при культивировании in vitro, придает постоянную способность к пролиферации,
хотя и с умеренной скоростью, в то время как удаление всех членов семейства Rb -генов делает клетки практически нечувствительными к ингибированию роста при культивировании in vitro и сильно увеличивает
скорость их размножения.
Immortal
TKO MEF сохраняют интактный путь p19 ARF /p53 Ранее предполагалось, что репликативное старение является результатом р53-зависимого ингибирующего эффекта роста
p19 ARF , который постепенно накапливается при длительном пассировании (Quelle et al. 1995; Zindy et al.1997). Таким образом, было показано, что инактивация Ink4a, p19 Arf или p53 приводит к иммортализации MEF (Serrano et al. 1996, 1997; Kamijo et al. 1997). Более того, спонтанная иммортализация MEF сопровождалась либо мутацией p53 (Harvey and Levine 1991; Rittling and Denhardt 1992), либо делецией локуса Ink4a (Kamb et al. 1994; Nobori et al. 1994; Zindy et al. ., 1998), удаляя как p16 INK4A , так и p19 ARF (Kamijo et al.1997). Поэтому мы проверили функциональность этого пути на бессмертных TKO MEF. На фигуре 5А показано, что p16 Ink4a и p19 Arf правильно экспрессировались во всех генотипах, включая TKO , и имели тенденцию к увеличению при длительном пассировании. Кроме того, функция р53 индуцировалась, о чем свидетельствовало повышение уровня белка и
накопление p21 CIP (фиг. 5Б). Кроме того, когда TKO MEF были подвергнуты гамма-облучению, они показали сильную индукцию p21 Cip , как и wt и Rb -/- p107 -/- MEFs, что снова указывает на то, что клетки содержат функциональный р53 (рис.5С). Эти наблюдения показывают, что полная потеря функции семейства генов Rb делает MEF устойчивыми к старению, индуцированному p19 ARF /p53, и предсказывают, что TKO MEF при количестве пассажей, когда MEF дикого типа спонтанно ускользают от старения, могут расти без мутации p53 или делеции локуса Ink4a .
На фигуре 5А показано, что p16 Ink4a и p19 Arf правильно экспрессировались во всех генотипах, включая TKO , и имели тенденцию к увеличению при длительном пассировании. Кроме того, функция р53 индуцировалась, о чем свидетельствовало повышение уровня белка и
накопление p21 CIP (фиг. 5Б). Кроме того, когда TKO MEF были подвергнуты гамма-облучению, они показали сильную индукцию p21 Cip , как и wt и Rb -/- p107 -/- MEFs, что снова указывает на то, что клетки содержат функциональный р53 (рис.5С). Эти наблюдения показывают, что полная потеря функции семейства генов Rb делает MEF устойчивыми к старению, индуцированному p19 ARF /p53, и предсказывают, что TKO MEF при количестве пассажей, когда MEF дикого типа спонтанно ускользают от старения, могут расти без мутации p53 или делеции локуса Ink4a . Действительно, мы обнаружили, что в шести независимых клонах, которые выращивались по протоколу на основе 3T9 в течение >20 пассажей, p19 ARF все еще экспрессировался, в отличие от клона wt 3T9, который утратил экспрессию p19 ARF , вероятно, из-за делеции локуса Ink4a (рис.5Д). Наконец, все шесть клонов TKO 3T9 показали повышенную экспрессию p21 Cip при γ-облучении, как и в wt и TKO MEF раннего пассажа, что указывает на то, что p53 также оставался функциональным в этих клонах (рис. 5E). .
Действительно, мы обнаружили, что в шести независимых клонах, которые выращивались по протоколу на основе 3T9 в течение >20 пассажей, p19 ARF все еще экспрессировался, в отличие от клона wt 3T9, который утратил экспрессию p19 ARF , вероятно, из-за делеции локуса Ink4a (рис.5Д). Наконец, все шесть клонов TKO 3T9 показали повышенную экспрессию p21 Cip при γ-облучении, как и в wt и TKO MEF раннего пассажа, что указывает на то, что p53 также оставался функциональным в этих клонах (рис. 5E). .
Интактный p19 Путь ARF /p53 в бессмертных TKO MEF.( A ) Указанные MEF пассировали в соответствии с протоколом 3T9, а осадки клеток собирали на пассажах 3, 5 и 7. Клетки
лизаты анализировали на уровни p16 INK4A , p19 ARF и тубулина с помощью иммуноблоттинга. Инк4а -/- MEF служили в качестве отрицательного контроля. ( B ) Экспрессия p53 и p21 CIP в wt и TKO MEF в пассажах 3 и 6.( C ) Анализ экспрессии p21 CIP в контроле и γ-облученных ранних пассажах wt , Rb -/- p107 -/- и TKO MEF, демонстрирующие индукцию p21 Cip у всех генотипов через 24 часа после облучения. ( D ) Анализ экспрессии p19 ARF в шести независимых клонах TKO 3T9 (1–6). Все клоны экспрессировали p19 Arf , тогда как клон 3T9 дикого типа (3T9 wt , 1) утратил экспрессию этого белка. стр.19 Арф -/- MEF служили в качестве отрицательного контроля. ( E ) Индукция p21 Cip в γ-облученных wt (1) и TKO (1–6) клеточных линиях 3T9 MEF по сравнению с γ-облученными клеточными линиями wt и TKO и TKO на ранних пассажах MEF.
Инк4а -/- MEF служили в качестве отрицательного контроля. ( B ) Экспрессия p53 и p21 CIP в wt и TKO MEF в пассажах 3 и 6.( C ) Анализ экспрессии p21 CIP в контроле и γ-облученных ранних пассажах wt , Rb -/- p107 -/- и TKO MEF, демонстрирующие индукцию p21 Cip у всех генотипов через 24 часа после облучения. ( D ) Анализ экспрессии p19 ARF в шести независимых клонах TKO 3T9 (1–6). Все клоны экспрессировали p19 Arf , тогда как клон 3T9 дикого типа (3T9 wt , 1) утратил экспрессию этого белка. стр.19 Арф -/- MEF служили в качестве отрицательного контроля. ( E ) Индукция p21 Cip в γ-облученных wt (1) и TKO (1–6) клеточных линиях 3T9 MEF по сравнению с γ-облученными клеточными линиями wt и TKO и TKO на ранних пассажах MEF.
p53 может индуцировать как pRB-зависимую G 1 , так и pRB-независимую G 2 /M остановку при гамма-облучении (Agarwal et al.1995; Бругаролас и др. 1995 год; Харрингтон и др. 1998). На рис. 6А показано, что вызванная γ-облучением остановка G 1 действительно была снята в Rb −/− p107 −/− и ТКО МЭФ, а блокпост Г 2 /М остался цел. Это еще раз указывает на то, что в руб. −/− p107 -/- и клеток TKO , p53 оставался функциональным, и что карманные белки необходимы для радиационно-индуцированной остановки G 2 /M.
Рисунок 6.
Дефектный Г 1 КПП в ТКО МЭФ на облучение и эктопию р19 Арф экспрессия. ( A ) γ-облучение вызвало остановку G 2 /M в Rb −/− p107 −/− и TKO MEF.Проценты G 1 , S и G 2 /M фаз клеточного цикла в необлученных и облученных (5,5 Гр) MEF были измерены с помощью анализа FITC-PI FACS и нанесены на график.
как точечные диаграммы FL3-A (ось X ) – FL1-H (ось Y ). ( B ) Эктопическая экспрессия p19 Arf индуцирует зависимую от карманного белка G 1 остановку. FACS-анализ GFP-позитивных Rb −/− , руб. −/− p107 -/- и TKO MEF, экспрессирующие векторную плазмиду, доминантно-негативный вариант CDK2, или p19 Arf (1. 0 или 5,0 мкг) вместе с конструкцией экспрессии гистона h3B-GFP в соотношении 10 : 1. Репрезентативные результаты трех независимых
показаны опыты.
0 или 5,0 мкг) вместе с конструкцией экспрессии гистона h3B-GFP в соотношении 10 : 1. Репрезентативные результаты трех независимых
показаны опыты.
Наконец, мы спросили, чувствительны ли MEF с дефицитом семейства Rb к остановке G 1 после эктопической экспрессии p19 ARF .Клетки подвергали электропорации и анализировали с помощью FACS. Во-первых, мы обнаружили, что введение CDK2 доминантно-негативного белка увеличивало
в г 1 населения на столько же в руб −/− , руб. −/− p107 -/- и TKO MEF (рис. 6B). Эктопическая экспрессия p19 ARF индуцировала подобную остановку G 1 в клетках, лишенных pRB или как pRB, так и p107. Напротив, в TKO MEF остановка G 1 лишь незначительно индуцировалась p19 ARF по сравнению с эффектом CDK2 dn (фиг. 6B). Вместе с наблюдением, что удаление семейства генов Rb вызывает иммортализацию, несмотря на нормальные уровни белков INK4A и функционального p53, этот результат указывает на то, что
карманные белки являются важными медиаторами реакции старения на p19 ARF .
Напротив, в TKO MEF остановка G 1 лишь незначительно индуцировалась p19 ARF по сравнению с эффектом CDK2 dn (фиг. 6B). Вместе с наблюдением, что удаление семейства генов Rb вызывает иммортализацию, несмотря на нормальные уровни белков INK4A и функционального p53, этот результат указывает на то, что
карманные белки являются важными медиаторами реакции старения на p19 ARF .
Инактивация карманных белков приводит к повышенному клеточному обороту в условиях ингибирования роста
Клетки TKO в значительной степени устойчивы к ингибированию роста p19 ARF .Поэтому мы исследовали поведение этих клеток в ряде других ингибирующих рост условий.
Истощение фактора роста
MEF были высеяны при низкой плотности, снабжены 0,1% сывороткой и проанализированы с помощью BrdU-FACS. Клетки Wt и Rb -/- сильно арестованы, о чем свидетельствует скопление клеток в G 1 и блокирование синтеза ДНК (рис.7А). Ответ руб. −/− p107 -/- МЭФ был менее выражен, хотя уровень синтеза ДНК все же значительно снижался по сравнению с 10% сывороткой (см.
также рис. 3А). Напротив, TKO MEF продолжали включать BrdU почти с той же скоростью, что и в условиях высокого содержания сыворотки. Удивительно, но в этих условиях
увеличения числа клеток при длительном культивировании не наблюдалось (рис.7Б). Появление плавающих клеток в среде через 24 ч после нанесения низкой сыворотки свидетельствовало о том, что клетки стали
апоптотический. Действительно, анализ PI-FACS выявил значительную суб-G 1 популяцию в TKO MEF через 24 часа после добавления низкой сыворотки, увеличивающуюся до 10% популяции через 4 дня (рис.
Клетки Wt и Rb -/- сильно арестованы, о чем свидетельствует скопление клеток в G 1 и блокирование синтеза ДНК (рис.7А). Ответ руб. −/− p107 -/- МЭФ был менее выражен, хотя уровень синтеза ДНК все же значительно снижался по сравнению с 10% сывороткой (см.
также рис. 3А). Напротив, TKO MEF продолжали включать BrdU почти с той же скоростью, что и в условиях высокого содержания сыворотки. Удивительно, но в этих условиях
увеличения числа клеток при длительном культивировании не наблюдалось (рис.7Б). Появление плавающих клеток в среде через 24 ч после нанесения низкой сыворотки свидетельствовало о том, что клетки стали
апоптотический. Действительно, анализ PI-FACS выявил значительную суб-G 1 популяцию в TKO MEF через 24 часа после добавления низкой сыворотки, увеличивающуюся до 10% популяции через 4 дня (рис. 7C). Опять же, руб. −/− p107 -/- показали промежуточный фенотип. Мы не наблюдали увеличения популяции многоядерных MEF, что указывает на то, что непрерывные
Включение BrdU не происходило в результате эндоредупликации (данные не показаны).
7C). Опять же, руб. −/− p107 -/- показали промежуточный фенотип. Мы не наблюдали увеличения популяции многоядерных MEF, что указывает на то, что непрерывные
Включение BrdU не происходило в результате эндоредупликации (данные не показаны).
Повышенный клеточный оборот в TKO MEF в условиях ингибирования роста. ( A ) Распределение фаз G 1 , S и G 2 /M указанных MEF при 0,1% FCS. ( B ) Кривые роста указанных MEF при 0,1% FCS. ( C ) Процент апоптотических клеток в указанных культурах MEF при 0.1% FCS, 24 ч (черный) и 96 ч (серый) после истощения факторов роста.
Проценты были получены путем определения популяции sub-G 1 на гистограмме FL3A-log. ( D ) Распределение фаз G 1 , S и G 2 /M указанных контактно-ингибируемых MEF. ( E ) Процент апоптотических клеток в контактно-ингибированных культурах MEF. Указаны средние значения трех независимых экспериментов.
в процентах.Столбики погрешностей указывают на стандартные отклонения от среднего значения ( A,C,D,E ).
( D ) Распределение фаз G 1 , S и G 2 /M указанных контактно-ингибируемых MEF. ( E ) Процент апоптотических клеток в контактно-ингибированных культурах MEF. Указаны средние значения трех независимых экспериментов.
в процентах.Столбики погрешностей указывают на стандартные отклонения от среднего значения ( A,C,D,E ).
Блокировка контактов
Другим внеклеточным сигналом, который провоцирует остановку G 0 /G 1 в нормальных фибробластах, является межклеточный контакт при слиянии (Nilausen and Green 1965).Было показано, что это явление, известное как контактное ингибирование, сопровождается повышением уровня p27 KIP1 и снижением уровня циклина D1 (Polyak et al. , 1994; St. Croix et al., 1998). Кроме того, накапливались гипофосфорилированные изоформы pRb, указывая на то, что pRB необходим для остановки G 1 . Для изучения роли карманных белков в контактном ингибировании MEF выращивали до слияния и культивировали в течение дополнительного времени.
4 д. Синтез ДНК измеряли с помощью BrdU-FACS. Опять же, делеция всех карманных белков выявила серьезный дефект в установлении
a G 0 /G 1 арест (рис.7Д). Вместо этого TKO MEF продолжали включать BrdU и генерировать апоптотические клетки (фиг. 7E).
, 1994; St. Croix et al., 1998). Кроме того, накапливались гипофосфорилированные изоформы pRb, указывая на то, что pRB необходим для остановки G 1 . Для изучения роли карманных белков в контактном ингибировании MEF выращивали до слияния и культивировали в течение дополнительного времени.
4 д. Синтез ДНК измеряли с помощью BrdU-FACS. Опять же, делеция всех карманных белков выявила серьезный дефект в установлении
a G 0 /G 1 арест (рис.7Д). Вместо этого TKO MEF продолжали включать BrdU и генерировать апоптотические клетки (фиг. 7E).
Условия неприлипания
Независимость от прикрепления является характеристикой большинства онкогенно трансформированных типов клеток и была приписана конститутивным
циклин E- (Fang et al. 1996) или A- (Guadagno et al. 1993) зависимая активность CDK2.В незлокачественных клетках остановка роста в неадгезивных условиях была связана с потерей циклина.
Активность E-CDK2 (Fang et al., 1996) и недофосфорилирование pRB и p107 (Schulze et al., 1996). Однако мы обнаружили, что TKO MEF, хотя и дают очень маленькие колонии, содержащие до 20 клеток, не способны к устойчивой пролиферации в мягком агаре.
(Таблица 1). Это указывает на то, что абляция активности карманных белков недостаточна для роста в неадгезивных условиях и предполагает
что потеря активности CDK2 может вызвать остановку роста, независимую от карманных белков.Эта точка зрения согласуется с нашим наблюдением
что клеток TKO все еще могут быть заблокированы удалением активности CDK2 (фиг. 6B). Зависимость от якоря TKO MEF согласуется с их неспособностью расти под кожей голых мышей (не показано), демонстрируя, что эти клетки
не трансформируется.
1993) зависимая активность CDK2.В незлокачественных клетках остановка роста в неадгезивных условиях была связана с потерей циклина.
Активность E-CDK2 (Fang et al., 1996) и недофосфорилирование pRB и p107 (Schulze et al., 1996). Однако мы обнаружили, что TKO MEF, хотя и дают очень маленькие колонии, содержащие до 20 клеток, не способны к устойчивой пролиферации в мягком агаре.
(Таблица 1). Это указывает на то, что абляция активности карманных белков недостаточна для роста в неадгезивных условиях и предполагает
что потеря активности CDK2 может вызвать остановку роста, независимую от карманных белков.Эта точка зрения согласуется с нашим наблюдением
что клеток TKO все еще могут быть заблокированы удалением активности CDK2 (фиг. 6B). Зависимость от якоря TKO MEF согласуется с их неспособностью расти под кожей голых мышей (не показано), демонстрируя, что эти клетки
не трансформируется.
Immortal TKO MEF не растут самостоятельно
В заключение, полная аблация семейства генов Rb не делает возможным рост, не зависящий от привязки; однако он серьезно нарушает остановку клеточного цикла в ответ на истощение факторов роста и контактное ингибирование.В этих ингибирующих рост условиях клетки с дефицитом карманного белка подвергаются повышенный оборот клеток.
Обсуждение
Мы создали линию клеток ЭС, несущих инактивирующие нарушения во всех трех членах семейства генов ретинобластомы, и обнаружили
что это сильно ограничивало их дифференцирующую способность. При MEF полная абляция семейства карманных белков серьезно
скомпрометирован G 1 контроль. Наиболее ярко это проявилось в обнаружении того, что MEF с тройным нокаутом были бессмертными и устойчивыми в непрерывном режиме.
пролиферация и апоптоз в условиях ограничения роста.
При MEF полная абляция семейства карманных белков серьезно
скомпрометирован G 1 контроль. Наиболее ярко это проявилось в обнаружении того, что MEF с тройным нокаутом были бессмертными и устойчивыми в непрерывном режиме.
пролиферация и апоптоз в условиях ограничения роста.
Дифференциация
ES-клеток имеют очень высокую скорость пролиферации (время удвоения 7–8 ч), а в пролиферирующей популяции около 70% клеток
были обнаружены в S-фазе (Savatier et al.1994). Саватье и др. (1996) сделали неожиданное наблюдение, что недифференцированные ЭС клетки не содержат обнаруживаемой активности циклина D1/CDK4 и являются
нечувствителен к ингибированию роста p16 INK4A . Более того, ES клетки не экспрессируют p130 (LeCouter et al. 1996), тогда как pRB и p107 могут быть легко обнаружены (Robanus-Maandag et al. 1998). По-видимому, в ЭС клетках антипролиферативные эффекты pRB и p107 нейтрализованы, возможно, наличием Е1А-подобного
активности (E1A-LA), как предполагалось ранее (Murray et al.1991). Резкое увеличение активности циклина D1/CDK4 при индукции дифференцировки, вероятно, отражает контроль pRB/p107/p130.
пути становятся активными. Наши наблюдения полностью согласуются с этой точкой зрения: абляция семейства pRB не влияла на
потенциал роста ЭС клеток, но серьезно нарушил их дифференцирующий потенциал. Таким образом, необходимы карманные белки.
для дифференцировки ЭС клеток. Более того, поскольку как ЭС клетки с дефицитом pRB/p107 (данная работа), так и ЭС клетки с дефицитом p130 (не
показан) показал нормальную мышечную и нейрональную дифференцировку при тератокарциномах, p130 может компенсировать отсутствие pRB
и стр.107.
1998). По-видимому, в ЭС клетках антипролиферативные эффекты pRB и p107 нейтрализованы, возможно, наличием Е1А-подобного
активности (E1A-LA), как предполагалось ранее (Murray et al.1991). Резкое увеличение активности циклина D1/CDK4 при индукции дифференцировки, вероятно, отражает контроль pRB/p107/p130.
пути становятся активными. Наши наблюдения полностью согласуются с этой точкой зрения: абляция семейства pRB не влияла на
потенциал роста ЭС клеток, но серьезно нарушил их дифференцирующий потенциал. Таким образом, необходимы карманные белки.
для дифференцировки ЭС клеток. Более того, поскольку как ЭС клетки с дефицитом pRB/p107 (данная работа), так и ЭС клетки с дефицитом p130 (не
показан) показал нормальную мышечную и нейрональную дифференцировку при тератокарциномах, p130 может компенсировать отсутствие pRB
и стр.107.
Несмотря на их ограниченную способность к дифференцировке при помещении под кожу бестимусных мышей, клетки TKO ES, введенные в бластоцисты, не демонстрировали неограниченную пролиферацию, но вместо этого были способны способствовать эмбриональному развитию. развитие по крайней мере до 15-го дня беременности, что позволяет получить ТКО МЭФ. Это наблюдение может свидетельствовать о том, что дифференцировка в определенные типы клеток, например, в фибробласты, не требует
карманно-белковая функция.Более интригующей является возможность того, что дифференцировка клеток TKO в химерных эмбрионах направляется и контролируется окружающими клетками дикого типа. В любом случае в процессе разработки
клетки, по-видимому, приобрели характеристики, которые, в отличие от их эмбриональных стволовых клеток-предшественников, препятствуют их росту.
закрепление самостоятельно или под кожей голых мышей.
развитие по крайней мере до 15-го дня беременности, что позволяет получить ТКО МЭФ. Это наблюдение может свидетельствовать о том, что дифференцировка в определенные типы клеток, например, в фибробласты, не требует
карманно-белковая функция.Более интригующей является возможность того, что дифференцировка клеток TKO в химерных эмбрионах направляется и контролируется окружающими клетками дикого типа. В любом случае в процессе разработки
клетки, по-видимому, приобрели характеристики, которые, в отличие от их эмбриональных стволовых клеток-предшественников, препятствуют их росту.
закрепление самостоятельно или под кожей голых мышей.
Увековечение
В первичных фибробластах человека и мыши длительное культивирование генерирует ингибирующий рост сигнал, который в конечном итоге приводит
к состоянию репликативного старения, отражаемого увеличенной, уплощенной морфологией и отсутствием синтеза ДНК (Hayflick and Moorhead 1961; Todaro and Green 1963; Campisi 1997). Этот тип остановки роста сопровождается постепенным повышением уровней ингибиторов CDK2/4, p21 CIP и p16 INK4A , ингибитора клеточного цикла p19 ARF и p53 (Lloyd et al., 1997; Palmero et al. 1997; Зинди и др., 1997, 1998). p16 INK4A и p19 ARF кодируются одним генетическим локусом, Ink4a , при этом p19 ARF экспрессируется из альтернативной рамки считывания (Quelle et al. 1995). В то время как p16 INK4A , как было показано, действует выше pRB, способствуя остановке клеточного цикла (Serrano et al.1993), p19 ARF может физически взаимодействовать с p53 и/или MDM2, тем самым противодействуя функции MDM2 и стабилизируя p53 (Kamijo et al. 1998; Pomerantz et al. 1998; Zhang et al. 1998). Спонтанная иммортализация MEF обычно сопровождается делецией локуса Ink4a (Kamb et al., 1994; Nobori et al., 1994; Kamijo et al., 1997; Zindy et al., 1998) или потерей функции p53 (Harvey and Levine).
Этот тип остановки роста сопровождается постепенным повышением уровней ингибиторов CDK2/4, p21 CIP и p16 INK4A , ингибитора клеточного цикла p19 ARF и p53 (Lloyd et al., 1997; Palmero et al. 1997; Зинди и др., 1997, 1998). p16 INK4A и p19 ARF кодируются одним генетическим локусом, Ink4a , при этом p19 ARF экспрессируется из альтернативной рамки считывания (Quelle et al. 1995). В то время как p16 INK4A , как было показано, действует выше pRB, способствуя остановке клеточного цикла (Serrano et al.1993), p19 ARF может физически взаимодействовать с p53 и/или MDM2, тем самым противодействуя функции MDM2 и стабилизируя p53 (Kamijo et al. 1998; Pomerantz et al. 1998; Zhang et al. 1998). Спонтанная иммортализация MEF обычно сопровождается делецией локуса Ink4a (Kamb et al., 1994; Nobori et al., 1994; Kamijo et al., 1997; Zindy et al., 1998) или потерей функции p53 (Harvey and Levine). 1991; Риттлинг и Денхардт, 1992). Кроме того, Ink4a −/− и стр.53 −/− MEF бессмертны.Эти наблюдения предполагают, что индукция белков INK4A и p53 причинно связана с индукцией
старение. Однако мыши и MEF, лишенные только p19 ARF , показали фенотип, неотличимый от фенотипа, возникающего в результате разрушения локуса Ink4a , отменяющего как p16 INK4A , так и p19 ARF . Хотя последствия удаления одного p16 INK4A неизвестны, это предполагает, что путь p19 ARF /p53, а не p16 INK4A /pRB, отвечает за индукцию старения.
1991; Риттлинг и Денхардт, 1992). Кроме того, Ink4a −/− и стр.53 −/− MEF бессмертны.Эти наблюдения предполагают, что индукция белков INK4A и p53 причинно связана с индукцией
старение. Однако мыши и MEF, лишенные только p19 ARF , показали фенотип, неотличимый от фенотипа, возникающего в результате разрушения локуса Ink4a , отменяющего как p16 INK4A , так и p19 ARF . Хотя последствия удаления одного p16 INK4A неизвестны, это предполагает, что путь p19 ARF /p53, а не p16 INK4A /pRB, отвечает за индукцию старения.
Теперь мы показываем, что MEF с дефицитом pRB, p107 и p130 бессмертны, не могут быть арестованы эктопической экспрессией p19 ARF и не поддерживают делеции локуса Ink4a или мутации в p53 при длительном пассировании. Кроме того, MEF с дефицитом pRB/p107 можно считать бессмертными: хотя они реагируют на супрафизиологические
уровни p19 ARF при аресте G 1 , после адаптации к культивированию in vitro они сохраняли умеренную, но постоянную скорость пролиферации при длительном
проход.Эти наблюдения показывают, что как старение, индуцированное p19 ARF , так и остановка клеточного цикла требуют членов семейства pRB. Таким образом, остается вероятным, что оба продукта
локус Ink4a участвует в индукции старения, p16 INK4A не является достаточным, но способствует активации карманных белков с помощью p19 ARF (Carnero et al. 2000). Как p19 ARF передает сигналы семейству pRB, остается неясным. Бессмертный фенотип p53 -/- MEFs предполагают, что этот путь зависит от p53.Однако p19 ARF -p53-индуцированное старение не реализуется через p21 CIP -индуцированное ингибирование циклина E/CDK2, как p21 −/− и стр.
Кроме того, MEF с дефицитом pRB/p107 можно считать бессмертными: хотя они реагируют на супрафизиологические
уровни p19 ARF при аресте G 1 , после адаптации к культивированию in vitro они сохраняли умеренную, но постоянную скорость пролиферации при длительном
проход.Эти наблюдения показывают, что как старение, индуцированное p19 ARF , так и остановка клеточного цикла требуют членов семейства pRB. Таким образом, остается вероятным, что оба продукта
локус Ink4a участвует в индукции старения, p16 INK4A не является достаточным, но способствует активации карманных белков с помощью p19 ARF (Carnero et al. 2000). Как p19 ARF передает сигналы семейству pRB, остается неясным. Бессмертный фенотип p53 -/- MEFs предполагают, что этот путь зависит от p53.Однако p19 ARF -p53-индуцированное старение не реализуется через p21 CIP -индуцированное ингибирование циклина E/CDK2, как p21 −/− и стр. 21 −/− стр.27 -/- MEF все еще подвергаются старению и/или реагируют на сверхэкспрессию p19 ARF (Pantoja and Serrano 1999; Groth et al. 2000). Кроме того, наши данные показывают, что TKO MEF не блокируются в G 1 сверхэкспрессией p19 ARF , хотя они все еще могут быть заблокированы ингибированием активности CDK2 при экспрессии доминантно-негативного
действующий мутант CDK2.Таким образом, возникает картина, в которой p19 ARF индуцирует старение через семейство pRB, но независимо от p21 CIP , опосредованного ингибированием активности циклина E/CDK2. Является ли связь между р19 ARF и семейством генов Rb зависимой от р53, остается неясным. В отличие от одного отчета (Kamijo et al. 1997), в другом показано, что p19 ARF может вызывать остановку клеточного цикла у p53 . -/- MEF, которые могут быть ослаблены гиперэкспрессией E2F-1 или блокировкой функции p16 INK4A (Carnero et al.
21 −/− стр.27 -/- MEF все еще подвергаются старению и/или реагируют на сверхэкспрессию p19 ARF (Pantoja and Serrano 1999; Groth et al. 2000). Кроме того, наши данные показывают, что TKO MEF не блокируются в G 1 сверхэкспрессией p19 ARF , хотя они все еще могут быть заблокированы ингибированием активности CDK2 при экспрессии доминантно-негативного
действующий мутант CDK2.Таким образом, возникает картина, в которой p19 ARF индуцирует старение через семейство pRB, но независимо от p21 CIP , опосредованного ингибированием активности циклина E/CDK2. Является ли связь между р19 ARF и семейством генов Rb зависимой от р53, остается неясным. В отличие от одного отчета (Kamijo et al. 1997), в другом показано, что p19 ARF может вызывать остановку клеточного цикла у p53 . -/- MEF, которые могут быть ослаблены гиперэкспрессией E2F-1 или блокировкой функции p16 INK4A (Carnero et al. 2000). Эти данные свидетельствуют о том, что p19 ARF может нацеливаться на путь pRB независимо от p53. Единственной известной мишенью p19 ARF является MDM2, который может связывать p53, но также и pRB (Xiao et al. 1995) и, таким образом, может быть кандидатом на опосредование соединения p19 ARF /pRB.
2000). Эти данные свидетельствуют о том, что p19 ARF может нацеливаться на путь pRB независимо от p53. Единственной известной мишенью p19 ARF является MDM2, который может связывать p53, но также и pRB (Xiao et al. 1995) и, таким образом, может быть кандидатом на опосредование соединения p19 ARF /pRB.
Роль карманных белков в онкогенной трансформации
Экспрессия так называемых трансформирующих онкогенов, таких как онкогенный RAS (Palmero et al.1998) и v-Abl (Radfar et al. 1998) сильно повышает уровень p19 ARF . Это активирует p53-зависимую контрольную точку, которая защищает клетки от аномальной митогенной передачи сигналов, индуцируя репликативную
арестовать. MEF, лишенные функции p19 ARF или p53, больше не восприимчивы к этому типу защиты; они избегают репликативного старения и становятся
онкогенно трансформируется (Harvey and Levine 1991; Harvey et al. 1993; Kamijo et al.1997). Было показано, что избыточная экспрессия другого класса онкогенов, таких как MYC, аденовирус E1A и E2F-1, в MEF усиливает пролиферацию.
но также и апоптоз (Lowe and Ruley 1993; Hermeking and Eick 1994; Wagner et al. 1994; Qin et al. 1994; Querido et al. 1997). Апоптоз значительно усиливается при лишении клеток внеклеточных факторов выживания (Evan et al., 1992; Lowe and Ruley, 1993) и зависит от функционального пути p19 ARF -p53, который не включает функцию p21 CIP (Lowe et al.1994 год; Вагнер и др. 1994 год; Бейтс и др. 1998 год; Де Станчина и др. 1998 год; Зинди и др. 1998). Поведение клеток, экспрессирующих MYC или E1A, очень напоминает поведение клеток с тройным нокаутом: абляция карманного белка
повышала пролиферативную способность клеток, но и их чувствительность к апоптозу: в частности, в условиях
низкая сыворотка или высокая плотность. Вырисовывается картина, что p19 ARF действует как сенсор аномальной или противоречивой митогенной передачи сигналов и активирует р53-зависимый ответ, который может вызвать
остановка клеточного цикла или сенсибилизация клеток к апотозу.
1993; Kamijo et al.1997). Было показано, что избыточная экспрессия другого класса онкогенов, таких как MYC, аденовирус E1A и E2F-1, в MEF усиливает пролиферацию.
но также и апоптоз (Lowe and Ruley 1993; Hermeking and Eick 1994; Wagner et al. 1994; Qin et al. 1994; Querido et al. 1997). Апоптоз значительно усиливается при лишении клеток внеклеточных факторов выживания (Evan et al., 1992; Lowe and Ruley, 1993) и зависит от функционального пути p19 ARF -p53, который не включает функцию p21 CIP (Lowe et al.1994 год; Вагнер и др. 1994 год; Бейтс и др. 1998 год; Де Станчина и др. 1998 год; Зинди и др. 1998). Поведение клеток, экспрессирующих MYC или E1A, очень напоминает поведение клеток с тройным нокаутом: абляция карманного белка
повышала пролиферативную способность клеток, но и их чувствительность к апоптозу: в частности, в условиях
низкая сыворотка или высокая плотность. Вырисовывается картина, что p19 ARF действует как сенсор аномальной или противоречивой митогенной передачи сигналов и активирует р53-зависимый ответ, который может вызвать
остановка клеточного цикла или сенсибилизация клеток к апотозу. Поведение клеток с тройным нокаутом указывает на то, что это решение зависит
на функции карманных белков. В их присутствии клетки останавливаются; при их отсутствии, например, путем генетической абляции, секвестрации
с помощью E1A или ингибирования после избыточной экспрессии MYC (Berns et al. 2000; Lasorella et al. 2000) клетки становятся бессмертными, но также очень чувствительными к апоптозу.
Поведение клеток с тройным нокаутом указывает на то, что это решение зависит
на функции карманных белков. В их присутствии клетки останавливаются; при их отсутствии, например, путем генетической абляции, секвестрации
с помощью E1A или ингибирования после избыточной экспрессии MYC (Berns et al. 2000; Lasorella et al. 2000) клетки становятся бессмертными, но также очень чувствительными к апоптозу.
Действительно, мы наблюдали, что культивирование MEF с тройным нокаутом в условиях, ограничивающих рост, легко индуцирует апоптоз.Однако в то же время наблюдалась продолжающаяся пролиферация клеток. Таким образом, при определенных условиях, ограничивающих рост,
потеря функции карманного белка обеспечивает непрерывный клеточный оборот без чистого увеличения числа клеток. Мы предвидим эту потерю
функции карманных белков in vivo может также привести к увеличению оборота клеток, что значительно облегчило бы приобретение
дополнительных онкогенных явлений. Несколько наблюдений поддерживают этот сценарий.Например, условная инактивация Rb в гипофизе мыши приводили к увеличению клеточного обмена. Опухоли, по-видимому, возникают из клеток, в которых происходит апоптоз.
был заблокирован, по-видимому, посредством дополнительной мутации (Vooijs et al. 1997). Кроме того, инактивация членов семейства pRB в эпителии головного мозга мыши большим Т-вариантом Т121 индуцировала аберрантные
пролиферацию и р53-зависимый апоптоз. Инактивация p53 в этих условиях привела к резкому снижению апоптоза.
(Инь и др.1997). Мы предполагаем, что избыточность карманных белков в контроле клеточного цикла зависит от типа клеток. Например, в мышке
гипофизе p107 и p130 могут не экспрессироваться или не функционировать, компенсируя потерю pRB, тогда как в сетчатке мыши
как pRB, так и p107 активны, и, следовательно, потеря обоих белков необходима для поддержания длительного обновления клеток в конечном итоге.
Несколько наблюдений поддерживают этот сценарий.Например, условная инактивация Rb в гипофизе мыши приводили к увеличению клеточного обмена. Опухоли, по-видимому, возникают из клеток, в которых происходит апоптоз.
был заблокирован, по-видимому, посредством дополнительной мутации (Vooijs et al. 1997). Кроме того, инактивация членов семейства pRB в эпителии головного мозга мыши большим Т-вариантом Т121 индуцировала аберрантные
пролиферацию и р53-зависимый апоптоз. Инактивация p53 в этих условиях привела к резкому снижению апоптоза.
(Инь и др.1997). Мы предполагаем, что избыточность карманных белков в контроле клеточного цикла зависит от типа клеток. Например, в мышке
гипофизе p107 и p130 могут не экспрессироваться или не функционировать, компенсируя потерю pRB, тогда как в сетчатке мыши
как pRB, так и p107 активны, и, следовательно, потеря обоих белков необходима для поддержания длительного обновления клеток в конечном итоге. приводящее к развитию ретинобластомы (Robanus-Maandag et al.1998).
приводящее к развитию ретинобластомы (Robanus-Maandag et al.1998).
В заключение мы определили три характеристики, связанные с потерей функции карманных белков, которые могут обеспечить обоснование его частого участия в раке человека. Это снижение способности к дифференцировке, потеря старения. реакция и повышенный оборот клеток в условиях, ограничивающих рост. Мы ожидаем, что тщательное изучение судьбы ЭС клетки, несущие одиночные или сложные мутации с потерей функции в семействе генов Rb у химерных мышей, позволят лучше понять роль абляции карманных белков в онкогенезе.
Материалы и методы
Получение ЭС клеток с дефицитом pRb, p107 и p130
Нокаутные аллели pRb, p107 и p130 были получены в эмбриональных стволовых клетках 129Ola (E14/IB10). Нацеливающие векторы 129Rb-hyg, 129Rb-his и 129p107-IRESβgeo
были описаны (te Riele et al.1992 год; Робанус-Маандаг и др. 1998). Для инактивации p130 мы создали нацеливающую конструкцию 129p130-pur, в которой кассета PGK-пуромицина была введена в кодон 405, находящийся
в А-домене области кармана (LeCouter et al. 1996). При электропорации и отборе при концентрации пуромицина 1,8 мкг/мл гомологичные рекомбинанты были получены с частотой 80%.
Оставшийся аллель p130 был инактивирован отбором при высоких концентрациях пуромицина (14 мкг/мл). с130 -/- мышей были созданы с использованием химерных мышей, дающих передачу по зародышевой линии.
Нацеливающие векторы 129Rb-hyg, 129Rb-his и 129p107-IRESβgeo
были описаны (te Riele et al.1992 год; Робанус-Маандаг и др. 1998). Для инактивации p130 мы создали нацеливающую конструкцию 129p130-pur, в которой кассета PGK-пуромицина была введена в кодон 405, находящийся
в А-домене области кармана (LeCouter et al. 1996). При электропорации и отборе при концентрации пуромицина 1,8 мкг/мл гомологичные рекомбинанты были получены с частотой 80%.
Оставшийся аллель p130 был инактивирован отбором при высоких концентрациях пуромицина (14 мкг/мл). с130 -/- мышей были созданы с использованием химерных мышей, дающих передачу по зародышевой линии.
Генерация MEF с дефицитом pRb, p107 и p130
химерных эмбриона получали путем инъекции мутантных ЭС клеток в бластоцисту C57Bl/6 возрастом 3,5 дня. Эмбрионов мы выделили 12
d после имплантации и введения в культуру.Вкратце, эмбриональную ткань без органов промывали в PBS, измельчали и инкубировали.
в 100 мкл трипсина/ЭДТА в течение ночи на льду, а затем 30 мин при 37°C с дополнительными 100 мкл трипсина/ЭДТА. Мутантные MEF были
выбран на 48 часов с 800 мкг/мл G418. Эти клетки были обозначены как пассаж 1. Нехимерные культуры MEF, служащие контролем,
все умерли в течение 48 часов после отбора.
Эмбрионов мы выделили 12
d после имплантации и введения в культуру.Вкратце, эмбриональную ткань без органов промывали в PBS, измельчали и инкубировали.
в 100 мкл трипсина/ЭДТА в течение ночи на льду, а затем 30 мин при 37°C с дополнительными 100 мкл трипсина/ЭДТА. Мутантные MEF были
выбран на 48 часов с 800 мкг/мл G418. Эти клетки были обозначены как пассаж 1. Нехимерные культуры MEF, служащие контролем,
все умерли в течение 48 часов после отбора.
Культура клеток ЭС, получение и характеристика тератокарцином
ЭС клеток культивировали, как описано (Robanus-Maandag et al.1998). Бестимусным мышам подкожно вводили 10 6 ЭС клеток. Опухоли удаляли через 4 недели после инъекции, фиксировали в фосфатно-буферном формалине, заливали в парафин, делали срезы. при 5 мкм и окрашивали гематоксилином и эозином по стандартным методикам. Для иммуногистохимической характеристики
регидратированные срезы окрашивали антителами против глиального фибриллярного кислого белка (GFAP, DAKO), низкоаффинного нервного
рецептор фактора роста (p75 LNGFR , Chemicon International), специфичную для нейронов энолазу (NSE, DAKO) и маркер пролиферации Ki-67 (MIB-5, Immunotech).Фракция
пролиферирующих клеток определяли путем подсчета Ki-67-позитивных клеток в шести различных областях среза тератокарциномы.
при увеличении 160х.
при 5 мкм и окрашивали гематоксилином и эозином по стандартным методикам. Для иммуногистохимической характеристики
регидратированные срезы окрашивали антителами против глиального фибриллярного кислого белка (GFAP, DAKO), низкоаффинного нервного
рецептор фактора роста (p75 LNGFR , Chemicon International), специфичную для нейронов энолазу (NSE, DAKO) и маркер пролиферации Ki-67 (MIB-5, Immunotech).Фракция
пролиферирующих клеток определяли путем подсчета Ki-67-позитивных клеток в шести различных областях среза тератокарциномы.
при увеличении 160х.
Культивирование MEF, кривые роста, протокол 3T9
MEF культивировали в GMEM (GIBCO) с добавлением 10% эмбриональной телячьей сыворотки, 1 мМ заменимых аминокислот, 1 мМ пирувата натрия,
и 0. 1 мМ β-меркаптоэтанол. Для кривых роста MEF высевали в количестве 2,5 × 10 4 клеток на 12-луночный планшет в трех экземплярах. В разные моменты времени клетки промывали PBS, фиксировали 5 мин в 4% растворе формальдегида.
и окрашивали 0,1% раствором кристаллического фиолетового (Sigma) в дистиллированной воде в течение 30 мин. После двукратного промывания клеток водой,
им дали высохнуть. Окрашивание клеток получали добавлением 1 мл 10% уксусной кислоты на лунку. Оптическая плотность была
измерено 100 мкл полученного окрашивания при 590 нм.Значения нормированы на оптическую плотность в день 0.
1 мМ β-меркаптоэтанол. Для кривых роста MEF высевали в количестве 2,5 × 10 4 клеток на 12-луночный планшет в трех экземплярах. В разные моменты времени клетки промывали PBS, фиксировали 5 мин в 4% растворе формальдегида.
и окрашивали 0,1% раствором кристаллического фиолетового (Sigma) в дистиллированной воде в течение 30 мин. После двукратного промывания клеток водой,
им дали высохнуть. Окрашивание клеток получали добавлением 1 мл 10% уксусной кислоты на лунку. Оптическая плотность была
измерено 100 мкл полученного окрашивания при 590 нм.Значения нормированы на оптическую плотность в день 0.
Протокол 3T9 выполняли путем посева 3 × 10 5 клеток на 6-луночный планшет в трех повторностях. Культуры инкубировали при 37°С в течение 3,5 сут и определяли общее количество клеток.
с использованием счетчика клеток (Casy 1) с последующим повторным посевом 3 × 10 5 клеток. Этот протокол был повторен 20 раз.
Этот протокол был повторен 20 раз.
Электропорация МЭФ
МЭФ выращивали в чашках диаметром 10 см до субконфлюэнтности.От одной до 2 × 10 6 клеток ресуспендировали в 100 мкл буфера для электропорации (2 мМ Hepes при pH 7,2, 15 мМ K 2 HPO 4 /KH 2 PO 4 9018 и 10 мМ 4 9218). мМ MgCl 2 ) при 37°С. Клеточную суспензию смешивали с плазмидной ДНК (экспрессирующей плазмидой гистона h3B-GFP [Kanda et al. 1998] вместе либо с доминантно-негативным вариантом CDK2 [Van den Heuvel and Harlow 1993], либо с pcDNA- p19 Arf . в соотношении 1 : 10) и переносили в кювету для электропорации (0.1 см, Биорад). Через 5 мин инкубации при комнатной температуре
клетки подвергали электропорации с помощью радиочастотного модуля Gene Pulser II/Gene Pulser II (Biorad). Через пять минут электропорация
клетки переносили в 8 мл среды при 37°C и высевали в 10-см чашку. Чтобы измерить влияние на G 1 , электропорированные клетки блокировали в G 2 /M нокодазолом (0,5 мкг/мл) через 32 часа после электропорации в течение 16 часов. Клетки трипсинизировали и ресуспендировали в 250 мкл 0.1%
Triton X-100 и 50 мкг/мл йодида пропидия в PBS. Для каждого образца с помощью FACS (FACscan,
Becton Dickinson) и проанализировали их распределение по клеточному циклу с помощью программы CellQuest (Becton Dickinson).
Через пять минут электропорация
клетки переносили в 8 мл среды при 37°C и высевали в 10-см чашку. Чтобы измерить влияние на G 1 , электропорированные клетки блокировали в G 2 /M нокодазолом (0,5 мкг/мл) через 32 часа после электропорации в течение 16 часов. Клетки трипсинизировали и ресуспендировали в 250 мкл 0.1%
Triton X-100 и 50 мкг/мл йодида пропидия в PBS. Для каждого образца с помощью FACS (FACscan,
Becton Dickinson) и проанализировали их распределение по клеточному циклу с помощью программы CellQuest (Becton Dickinson).
Анализ FACS
Распределение MEF в клеточном цикле определяли путем инкубации MEF с 5-бромдезоксиуридином (0.3 мкг/мл) плюс 5-фтор-5-дезоксиуридин
(0,03 мкг/мл) в течение 5 часов. Клетки собирали и ресуспендировали в 70% спирте при 4°C, окрашивали мышиным антителом DAKO против BrdU. и подготовлены для анализа FACS, как описано в Brugarolas et al. (1995).
и подготовлены для анализа FACS, как описано в Brugarolas et al. (1995).
Для анализа размера клеток культуры трипсинизировали, фиксировали в 70% спирте при 4°C, окрашивали йодидом пропидия и анализировали. с помощью FACS, как описано у Brugarolas et al.(1998).
Анализ белка
Уровни белка определяли вестерн-блоттингом в соответствии с установленными протоколами. Антитела против pRB (C-15), p107 (C-18), p130 (C-20), p16 INK4A (M-156), CDK4 (C-22), циклин E (M-20), p21 CIP (C-19) и p53 (FL-393) были получены от Санта-Крус; антитело p19 ARF (R562) было получено от Abcam.Козьи антикроличьи и козьи антимышиные вторичные антитела были получены от Biosource.
Облучение МЭФ
После того, как 5 × 10 5 клеток были высеяны на 10-сантиметровые чашки, их на следующий день облучили γ-излучением в дозе 20 Гр. Клеточные лизаты готовили
через 24 ч и анализировали на Вестерн-блоттинге.
Клеточные лизаты готовили
через 24 ч и анализировали на Вестерн-блоттинге.
Для анализа клеточного цикла клетки облучали 5.5 Гр. Через 14 ч клетки инкубировали с BrdU/FdU в течение 4 ч и анализировали. по ФАКС.
Анализ на мягком агаре
В GMEM, содержащем 10% сыворотки, 3 × 10 4 клеток ресуспендировали в 2 мл 0,4% агарозы с низкой температурой плавления (Sigma) и высевали в двух повторностях в шестилуночные планшеты, покрытые с 1% агарозы с низкой температурой плавления в GMEM, содержащей 10% сыворотки.Количество очагов подсчитывали через 2 нед.
Благодарности
Мы благодарим Karin van ‘t Wout и Karin van Veen-Buurman за инъекции бластоцисты и инъекции ES-клеток голым мышам;
Эльс Робанус-Маандаг и Марлин Деккер за предоставление рублей −/− ES-клетки и векторы-мишени; Kees de Goeij, Dennis Hoogervorst и Jurjen Bulthuis за гистотехническую помощь; Мартин
ван дер Валк для гистологического исследования тератокарциномы; Thijn Brummelkamp для экспрессионной плазмиды pcDNA-p19 ARF ; Чарльз Шерр и Фредерик Зинди за стр. 19 Арф −/− MEF; Джос Джонкерс для p53 −/− MEF; Рональд ДеПиньо для Ink4a −/− MEF; Эрик Нотебум и Анита Пфаут за помощь в анализе FACS; и Реувен Агами, Даниэль Пипер и Рене Бернар для
полезные обсуждения и критическое прочтение рукописи.Работа выполнена при поддержке Нидерландского онкологического фонда.
через проектный грант NKI 95–956 для H.t.R. (Дж.-Х.Д., А.в.Р., Л.С.).
19 Арф −/− MEF; Джос Джонкерс для p53 −/− MEF; Рональд ДеПиньо для Ink4a −/− MEF; Эрик Нотебум и Анита Пфаут за помощь в анализе FACS; и Реувен Агами, Даниэль Пипер и Рене Бернар для
полезные обсуждения и критическое прочтение рукописи.Работа выполнена при поддержке Нидерландского онкологического фонда.
через проектный грант NKI 95–956 для H.t.R. (Дж.-Х.Д., А.в.Р., Л.С.).
Затраты на публикацию этой статьи были частично покрыты за счет оплаты страниц. Таким образом, эта статья должна быть настоящим помечены как «реклама» в соответствии с разделом 1734 18 USC исключительно для обозначения этого факта.
Сноски
↵1 Автор, ответственный за переписку.

ЭЛЕКТРОННАЯ ПОЧТА hriele{at}nki.nl; ФАКС 31-20-512-2086.
Статья и публикация находятся на сайте www.genesdev.org/cgi/doi/10.1101/gad.847700.
- Поступила в редакцию 1 сентября 2000 г.
- Принят 25 октября 2000 г.
- Лабораторный пресс в Колд-Спринг-Харбор
Wind Cave NP: Путеводитель (1940)
| ПЕЩЕРА ВЕТРОВ Путеводитель 1940 г.  |
| ОТКРЫТЫЙ ВСЕ ГОД | | | Пещера Винд НАЦИОНАЛЬНЫЙ ПАРК ЮЖНАЯ ДАКОТА | |
| ВХОД В ПАРК С ЗАПАДА | ||||
Район Блэк-Хиллз в Южной Дакоте, где Ветер
Пещерный национальный парк имеет захватывающую историю создания земли.
рассказать.Интерес к ней варьируется от древних отложений окаменелостей, погребенных в
Бесплодные земли, рассказывающие историю о доисторических аллигаторах, носорогах,
трехпалых лошадей, и других давно вымерших животных, к высокой «игле»
образования, которые эрозия вылепила из массивов гранита высоко
покрытые лесом склоны Черных Холмов.
Пещера Ветра находится в великом известняке Пахасапа. формация, которая также содержит несколько других крупных подземных пещер. Факты об открытии пещеры малоизвестны, но это обычно считается, что он был открыт Томом Бингхэмом, уроженцем Блэк-Хиллз. пионером во время охоты на оленей в 1881 году.Его привлек странный насвистывая и пошарив в подлеске, он обнаружил что это было вызвано ветром, убегающим через небольшое отверстие в некоторых скалах. Эта дыра, не более 10 дюймов в диаметре, является единственным естественным вход в пещеру до сих пор обнаружен. Он находится в нескольких шагах позади нынешний вход в пещеру.
Сильные потоки ветра, которые дуют попеременно в
и из устья пещеры предложил свое имя. Это странно
явление, как полагают, вызвано изменениями в атмосферном
давление снаружи.Когда барометр падает, обычно дует ветер
наружу; когда он поднимается, дует ветер. Многим посетителям нравится останавливаться
у входа в пещеру вывешивать себя по погоде.
Нынешнее отверстие в пещере было сделано путем раскопок около 6 футов до длинной извилистой трещины или туннеля, ведущего в коридоры и галереи украшены разнообразными кристаллическими образованиями. Эти образования радикально отличаются от тех, что встречаются в большинстве пещер. потому что сталактитов и сталагмитов здесь практически нет. пещера, особенность, которая добавляет ей уникальности.Здесь образования относятся к необычному коробчатому и морозостойкому типу. Коробка состоит из нежно окрашенных кристаллов, расположенных в виде сот. Крошечный белый кристаллы, иногда накладывающиеся на розовый фон, висят гроздьями с потолков и карнизов, образуя редкостной красоты морозное украшение. Протяженность пещеры составляет примерно 10 миль, но она не была полностью исследована. исследовал.
Национальный парк Винд-Кейв был создан актом
Конгресс от 9 января 1903 г.Его границы были впоследствии
расширен, и теперь он включает в себя площадь 11 818,94 акра. Как и все остальные
национальные парки это заповедник дикой природы. буйвол, лось, антилопа и
олени обитают в этом районе, и их часто можно увидеть с главной дороги.
Как и все остальные
национальные парки это заповедник дикой природы. буйвол, лось, антилопа и
олени обитают в этом районе, и их часто можно увидеть с главной дороги.
ВХОД В ВЕТРОВУЮ ПЕЩЕРУ
КРАСОЧНАЯ ИСТОРИЯ
В дополнение к своей естественной красоте и научному интерес, юго-западная часть Южной Дакоты имеет красочный и живописная история.Возможно, французский исследователь пробрался туда как еще в 1683 году, и, как говорят, его посетили братья Верендри. в 1743 г.
Индейцы сиу, племя заметное даже среди
Индейцы за силу и отвагу давно оккупировали этот регион и только
подчиняется белому поселению после ожесточенной и трагической борьбы. Этот
племя, как полагают, произошло к востоку от Alleghenies, но как
уже в 1632 году французы нашли их в Висконсине и Миннесоте.Некоторые из
их потомки сегодня живут на Пайн-Ридж и в индейском районе Роузбад.
Бронирование, в нескольких минутах езды от парка Wind Cave.
Согласно индийской легенде, четыре ветра были главных божеств равнинных племен, а ветер ассоциировался в их вере с дыханием жизни и жизненным принцип. Поэтому Пещера Ветров была для них священным местом. Многие У племен Равнин были мифы, в которых рассказывалась история о том, как буйвол впервые вышел из пещеры (это была объективация тот факт, что всякая плоть сделана из земли), и вождь Джозеф Белый Бык (Pte Сан Хунка), живущий племянник Сидящего Быка, сказал, что Сиу верили, что Пещера Ветра в Блэк-Хилс была пещерой откуда Вакан Танка, Великая Тайна, отправила их в сиу Охотничьи угодья.Это была одна из причин, почему сиу так упорно боролись за Блэк-Хиллз, когда они были захвачены белыми. У начальника тоже есть сказал, что некоторые из его людей все еще надеялись, что, когда они вернут себе милости своих богов буйволы снова выйдут из этой пещеры и заполнить равнины. 1
1 «Тропа войны», Стэнли Вестал.

Тучи войны, закончившейся конфликтом 1861-65 гг. и индейские восстания под предводительством Красного Облака, Сидящего Быка, Пятнистый Хвост и другие великие воины мешали развитию этого области, но в 1874 году экспедиция через Блэк-Хиллс под руководством ген.Джордж А. Кастер привел к открытию золота на Френч-Крик и заселение последовало быстро. С 1879 по 1886 год были дни бума, которые сделали легендарные фигуры таких первопроходцев, как Дикий Билл Хикок, Дедвуд Дик, Бедствие Джейн и Проповедник Смит. Все четверо похоронены в Дедвуд.
Задолго до того, как Черные Холмы были известны
живописной красоты, которая привела к созданию Государственного парка Кастер в
сердце этого труднопроходимого региона, и строительство было начато на
Мемориал Рашмора, регион был известен во всем мире своими
полезные ископаемые, особенно золото.Шахта Хоумстейк в Лиде — это
Крупнейший золотой рудник в США. В 1933 году почти четверть
Золотодобыча США происходила из Черного
Холмы. С момента начала добычи в 1875 г. общее количество золота составило около
В этом регионе было добыто 300 000 000 долларов. Некоторые из других
полезных ископаемых, найденных в регионе, представляющих экономическую ценность, — это серебро, свинец,
медные, железные, оловянные и вольфрамовые руды; колумбит, танталит, слюда,
минералы мышьяка, лития и цезия; фуллерова земля, бентонит,
вулканический пепел, уголь, нефть и конструкционные материалы.
С момента начала добычи в 1875 г. общее количество золота составило около
В этом регионе было добыто 300 000 000 долларов. Некоторые из других
полезных ископаемых, найденных в регионе, представляющих экономическую ценность, — это серебро, свинец,
медные, железные, оловянные и вольфрамовые руды; колумбит, танталит, слюда,
минералы мышьяка, лития и цезия; фуллерова земля, бентонит,
вулканический пепел, уголь, нефть и конструкционные материалы.
| РАЗРЕЗ ПО ВОСТОЧНОМУ СКЛОНУ ЧЕРНЫХ ХОЛМОВ ПОДЪЕМ, ЧЕРЕЗ ВЕТРОВУЮ ПЕЩЕРУ, ВЗГЛЯД НА СЕВЕР |
ГЕОЛОГИЧЕСКАЯ ИСТОРИЯ ВЕТРОВОЙ ПЕЩЕРЫ
Пещеры в целом можно разделить на три
деления в зависимости от породы, в которой они образовались.
Они могут образоваться в известняке, магматических породах или песчанике. Из
эти известняковые пещеры являются наиболее важными и наиболее часто посещаемыми.
большой размер. Пещера Ветра представляет собой известняковую пещеру.
Пещера Ветра представляет собой известняковую пещеру.
Слой известняка, в котором образовалась Пещера Ветра. Мощность варьируется в районе Блэк-Хиллз от 300 до 630 футов. Это известен как известняк Пахасапа, местное образование в Миссисипи. возраст отложился в большом внутреннем море около 300 миллионов лет назад. Следующий отложения этого известняка, он был поднят над морем. Несколько таких периодов подъема и опускания произошло во время геологическая история.В эти периоды погружения Пахасапа известняк был перекрыт другими отложениями на несколько сотен футов в глубину. толщина. Окончательный взброс суши, вероятно, произошел во время последняя часть мелового периода, около 60 миллионов лет назад. То начало образования Пещеры Ветра, вероятно, датируется этим время.
Известняк, подвергшийся нескольким таким
периоды поднятия или искривления становятся трещиноватыми или сломанными. Эти трещины
развиваться во всех направлениях и под всеми углами. Близкие вместе служили
как шаблон для характерной «коробки», сформированной в следующих
манера: дождевая вода, просачивающаяся с поверхности через разлагающиеся
растительность поглощает углекислый газ, что делает воду более растворимой
для известняка. Эта содержащая углекислый газ вода уносит часть известняка в
раствора, затем при испарении откладывает его в трещины или щели
ниже. Позже более растворимый известняк между ребрами коробки был
растворился, оставив форму коробки такой, какой она видна сегодня.
Близкие вместе служили
как шаблон для характерной «коробки», сформированной в следующих
манера: дождевая вода, просачивающаяся с поверхности через разлагающиеся
растительность поглощает углекислый газ, что делает воду более растворимой
для известняка. Эта содержащая углекислый газ вода уносит часть известняка в
раствора, затем при испарении откладывает его в трещины или щели
ниже. Позже более растворимый известняк между ребрами коробки был
растворился, оставив форму коробки такой, какой она видна сегодня.
ТИПОВОЙ ПРИМЕР ФОРМИРОВАНИЯ КОРОБКИ
ДИКАЯ ЖИЗНЬ
Еще одна новая достопримечательность в Национальном парке Пещеры Ветра. представляет собой демонстрацию представителя дикой природы того типа дичи, которая бродила холмы Дакота в первые дни. Защищенный здесь от охотника, путешественник найдет большое стадо бизонов и небольшие группы антилоп, лось и олень.
Посетители обычно могут получить близкий, беспрепятственный вид
стада бизонов с главной магистрали, которая вьется по холмистой местности
холмы парка. Вся территория парка огорожена забором, который
удерживает животных от блуждания по диапазону. Это следует помнить
что игра дикая и с ней нельзя обращаться как с домашними животными. это
поэтому неразумно выходить из машины или приближаться к любому из
игровые животные.
Вся территория парка огорожена забором, который
удерживает животных от блуждания по диапазону. Это следует помнить
что игра дикая и с ней нельзя обращаться как с домашними животными. это
поэтому неразумно выходить из машины или приближаться к любому из
игровые животные.
В этом парке вся дикая природа представлена в естественном виде. среда обитания, давая зрителю острые ощущения от наблюдения за дикими животными в открыто, а не из-за заборов или решеток.
Небольшие группы антилоп часто можно увидеть на внимательный наблюдатель. Лоси часто посещают открытые части парка во время ночь, поздний вечер и раннее утро. В течение дня они вернуться к бревенчатым участкам. Олень обычно остаются в лесу, но иногда их можно увидеть пересекающими луга.
КАРТА ПОДЗЕМНЫХ МАРШРУТОВ — НАЦИОНАЛЬНЫЙ ПАРК ПЕЩЕРА ВЕТРОВ
(нажмите на изображение, чтобы открыть PDF-версию)
ШТАБ-КВАРТИРА ПАРКА, РАСПОЛОЖЕННАЯ СРЕДИ РОЛЛИНГ-ХИЛС
ОБЩАЯ ИНФОРМАЦИЯ
Национальный парк Винд-Кейв находится в ведении
Служба национальных парков Министерства внутренних дел США,
и представителем этого бюро, отвечающего за парк, является Гарри Дж. Лиек, суперинтендант. Его адрес: Хот-Спрингс, С. Дак.
Лиек, суперинтендант. Его адрес: Хот-Спрингс, С. Дак.
ПУТЕШЕСТВИЕ В ПЕЩЕРУ.— Все путешествия по пещере запрещены. под руководством компетентных смотрителей парка. Входные билеты такие следует:
| | центов | |
| 71541 | 75 | 95 |
| детей, 12-16 лет | 25 | |
| детей, 5 до 12 лет | 15 | |
| детей до 5 лет допустили бесплатно. | ||
Этот прейскурант включает использование лифта на выездная поездка.
Путешествие по пещере мало чем отличается от обычного
поход по горной тропе. Следует носить удобную обувь для ходьбы.
Температура 47° и не меняется зимой или летом. это
желательно взять легкий свитер или куртку. Пещера электрически
освещенный.
ОБСЛУЖИВАНИЕ ЛИФТОВ.—В 1935 г. для удобства посетителей в пещере был установлен лифт.Это избавляет от необходимости вылезать из пещеры.
ОБРАЗЦЫ ОБРАЗЦОВ. Образцы образований из пещера выставлена в здании штаб-квартиры для изучения и общий осмотр со стороны общественности. Обращение с формациями пещеры или сбор сувениров строго запрещен.
Летом также проводится выставка полевых цветов. поддерживается. Он включает в себя все распространенные виды флоры Блэк-Хиллз для местность.Сбор цветов в парке без разрешения администрации суперинтенданта строго запрещено.
РАЗМЕЩЕНИЕ. — Нет отелей или туристических домиков. расположены в парке, а общественный палаточный лагерь с бесплатными дровами и вода поддерживается возле штаб-квартиры. Столовая и предоставляются услуги газировки. Товары для отдыхающих, диковинки и различные статьи доступны.
Горячие источники — южный вход в Черное море. Холмы.Это в 10 милях к югу от национального парка Винд-Кейв в США.
Шоссе № 85А. Это почтовое отделение парка и хорошие отели.
лагеря и гаражи. В Хот-Спрингс есть национальный санаторий и государственный
солдатский дом и известен как живописный город Черного
Холмы.
Холмы.Это в 10 милях к югу от национального парка Винд-Кейв в США.
Шоссе № 85А. Это почтовое отделение парка и хорошие отели.
лагеря и гаражи. В Хот-Спрингс есть национальный санаторий и государственный
солдатский дом и известен как живописный город Черного
Холмы.
СТАДА ЗУБРОВ ЧАСТО ВИДЕЛИ С ШОССЕ
КАК ДОБРАТЬСЯ В ПАРК
НА АВТОМОБИЛЕ. Национальный парк Пещеры Ветра открыт. Шоссе США No.85A, и до него можно добраться боковыми рейсами из либо по шоссе № 16 США в Кастере, либо по шоссе № 1. 87 через Государственный парк Кастер.
ПО ЖЕЛЕЗНОЙ ДОРОГЕ.—До парка можно добраться с
следующие железнодорожные станции: Хот-Спрингс, Чикаго, Берлингтон и
Куинси и Чикагские и Северо-Западные железные дороги; Прингл и
Кастер на железной дороге Чикаго, Берлингтона и Куинси; и
Баффало-Гэп, Чикаго и Северо-Запад. Пассажиры
желающие посетить Национальный парк Винд-Кейв в качестве дополнительной поездки из Эджмонта или
Buffalo Gap (28 и 14 миль соответственно от Хот-Спрингс) может остановиться
закончились экскурсионные билеты. Из многих восточных пунктов билеты в
Национальные парки Йеллоустоун или Глейшер хороши для прохождения
Эджмонт.
Из многих восточных пунктов билеты в
Национальные парки Йеллоустоун или Глейшер хороши для прохождения
Эджмонт.
НА АВТОБУСЕ. Хот-Спрингс обслуживается компанией Black Hills. Транспортная компания из Шайенна, штат Вайоминг, на юг через Рапид. Город, С. Дак. В Шайенне установлены связи с Берлингтоном. Транспортная компания и межгосударственные транзитные линии. С востока и До северных Хот-Спрингс можно добраться через Рапид-Сити через Рапид-Сити. Линии, линии Red Ball и автобусные линии Palace City от Huron, Mitchell, и Су-Фолс, С.Дак. Из Бисмарка, Северная Дакота, центральная автобусная линия обслуживает Пьера, S. Dak., соединяющийся с линиями Rapid City Lines для Rapid City и Hot. Пружины.
НА САМОЛЕТЕ.— Высокоскоростные роскошные самолеты.
со всех точек до Шайенна можно добраться через United Air Lines.
Wyoming Air Service, выполняющая рейсы из Пуэбло, штат Колорадо, в Биллингс, штат Монтана,
также останавливается в Шайенне.
МОСТ НА ТРОП, ВЕДУЩИЙ К ВХОДУ В ПЕЩЕРУ
ДОСТОПРИМЕЧАТЕЛЬНОСТИ
Рейнджеры на стойке информации парка помогут в планирование поездки по Черным Холмам, а также дать информацию о другие национальные парки.В путешествии около 300 миль много интересных мест, включая Башню Дьявола, Пещеру Драгоценностей, Национальные памятники Fossil Cycad, Scotts Bluff и Badlands, Кастер Государственный парк, три национальных леса, федеральные и государственные рыбоводные заводы, а также можно посетить птичий заповедник.
НАЦИОНАЛЬНЫЙ ПАМЯТНИК ДРАГОЦЕННАЯ ПЕЩЕРА, переданный Службе национальных парков из Лесная служба Указом Президента от 1 апреля 1934 г. открыт для публики в июне, июле и августе.Это 14 миль к западу от Кастера по шоссе 16. В пещере есть ряд переходы и небольшие помещения, стены которых облицованы кальцитом кристаллы.
НАЦИОНАЛЬНЫЙ ПАМЯТНИК ИСКОПАЕМОГО ЦИКАДА, созданный в 1922 году для сохранения крупных залежей ископаемых
остатки папоротниковидных растений, которые росли много миллионов лет назад в
Мезозойский период, расположен на южной окраине Блэк-Хиллз,
недалеко от Пещеры Ветра. Памятник находится в нескольких милях от г.
шоссе и не доступен для автомобилей, кроме как через частную собственность.Поскольку этот район представляет интерес в основном для палеоботаников и геологов,
в настоящее время не находится в открытом доступе. Хороший образец
интересная окаменелость выставлена в национальном парке Wind Cave.
штаб-квартира.
Памятник находится в нескольких милях от г.
шоссе и не доступен для автомобилей, кроме как через частную собственность.Поскольку этот район представляет интерес в основном для палеоботаников и геологов,
в настоящее время не находится в открытом доступе. Хороший образец
интересная окаменелость выставлена в национальном парке Wind Cave.
штаб-квартира.
НАЦИОНАЛЬНЫЙ ПАМЯТНИК БЕДРОДОВ.— Резкий контраст в зеленой стране Блэк-Хилс находятся бесплодные земли Уайт-Ривер, бесплодная безлесная местность недалеко к юго-востоку от Пещеры Ветра. Здесь природа украсила землю всеми оттенками желтовато-коричневого, кремового, бледно-зеленый, золотой и розовый.Фантастически вырезанные эрозионные формы возвышаются над долинами, некоторые из них высотой от 150 до 300 футов. постоянно меняющийся цвет и странные образования делают этот регион сильным образное обращение.
БАШНЯ ДЬЯВОЛА НАЦИОНАЛЬНЫЙ ПАМЯТНИК. Еще один необычный
природный феномен страны Черных Холмов – Башня Дьявола
через границу штата в Вайоминге. Это большая масса изверженной породы
возвышается на 600 футов над рекой Бель-Фурш.
Это большая масса изверженной породы
возвышается на 600 футов над рекой Бель-Фурш.
АДМИНИСТРАТИВНОЕ ЗДАНИЕ Грант фото
ГЛОССАРИЙ
Краткое описание технических терминов, используемых в этот буклет и посетители через направляющую пещеру рейнджеров:
АРАКОНИТ (4 1 ) — Минерал, который представляет собой одну из форм карбоната кальция (CaCO 3 ), белого или окрашенного, который часто встречается в виде составных или излучающих групп кристаллов.
BOXWORK (3 1 ) — уникальные соты образование пещеры Ветра, состоящей из кальцита с небольшим количеством гематит.
КАЛЬЦИТ (3 1 ) — Минерал, представляющий
наиболее распространенная форма карбоната кальция, которая вскипает в кислоте.
Образования в известняковых кавернах состоят в основном из
кальцит.
CHERT (7 1 ) — очень твердый, аморфный форма кремнезема, которая часто встречается вместе с известняком.В Пещера Ветра кремень содержит ископаемые породы.
КОНКРЕТ — Округлый агрегат минерального вещество, образованное осаждением или отложением вокруг некоторого ядра (не образование пещеры).
КОНГЛОМЕРАТ — осадочная горная порода, состоящая в основном из сцементированного окатаного гравия.
МЕЛОВОЙ — последний период мезозоя. эпоха.
КАПЕЛЬНИК (3 1 ) — неправильной формы отложения карбоната кальция, выпадающие в осадок при испарении вода, просачивающаяся сквозь стены пещеры; сталактиты и сталагмиты иногда называют капельным камнем.
ERA — основное подразделение зарегистрированных геологических время.
РАЗЛОМ — Смещение или движение горных массивов. по плоскости излома,
FLOWSTONE (3 1 ) — карбонат кальция
(CaCO 3 ) отложения, образующиеся в результате осаждения из капельной или проточной воды
в основном по стенам.
ИСКОПАЕМЫЕ Остатки или следы древних животные или растения, сохранившиеся в осадочных породах, такие как раковины или следы животных или птиц.
FROSTWORK (3 1 ) — хрупкий заполнитель кристаллов кальцита, напоминающих иней; вообще чисто белый.
МАГНИТНЫЕ ПОРОДЫ — Горные породы, образованные охлаждение и затвердевание расплавленного каменного материала.
ИЗВЕСТНЯК (4 1 ) — осадочная горная порода. состоит в основном из карбоната кальция. Он может быть произведен компанией действием водорослей и беспозвоночных или осаждением кальция карбонат из воды.
МЕТАМОРФИЧЕСКИЕ ПОРОДЫ — магматические или осадочные породы изменяется под воздействием тепла и давления. Сланец представляет собой метаморфизованный сланец, а мрамор метаморфизованный известняк.
МИССИСИПИАН — период палеозойской эры.
ОБНАЖЕНИЕ — обнажение породы на поверхности.
ПАЛЕОЗОЙ — третья великая эра письменных геологическое время. Время большого развития беспозвоночных, рыб, и папоротниковидные деревья. Эпоха обычно подразделяется на семь периодов: Кембрийский (самый старый), ордовикский, силурийский, девонский, миссисипский, Пенсильванский и Пермский.
ОСАДОЧНЫЕ ПОРОДЫ — Породы, образованные накоплением осадок в воде или на суше; может состоять из сланца, известняка, песчаник, окаменелости, гипс или лёсс.
СТАЛАКТИТЫ — известняковые конусы, свисающие с крыши известняковых пещер и образованы известковыми водами которые просачиваются через крышу.
СТАЛАГМИТЫ — Структуры, подобные сталактитам, которые развиваются на дне известняковых пещер и растут вверх за счет приростов от воды, капающей на них с потолка пещеры.Обычно больше тупее, чем сталактиты.
ТРАВЕРТИН (3 1 )——кальций
карбонат (CaCO 3 ), осажденный из раствора родниками или
проточная вода. Охватывает многие виды отложений.
Охватывает многие виды отложений.
Шкала твердости минералов:
1-тальк
2-гипс
3-кальцит
4-флюорит
5-Апатит
6-Ортоклаз
7-кварц
8-Топаз
9-Корунд
10-ромб
ПРАВИЛА И ПОСТАНОВЛЕНИЯ
ПРАВИЛА ПАРКА предназначены для охраны природных красот и пейзажей, а также для комфорта и удобство посетителей.Следующий синопсис предназначен для общего руководство посетителями, которых просят помочь администрации соблюдая правила. С полным регламентом можно ознакомиться в офисе суперинтендант и рейнджер станции.
УСЛУГИ ГИДОВ.— Никому не разрешается входить пещера, если только в сопровождении суперинтенданта или другого парка наемный рабочий.
ПОЖАРЫ. Зажгите осторожно и в специально отведенных местах.
места. Полностью погасить перед выходом из лагеря, даже при временном отсутствии. Делать
не догадывайся, что твой огонь погас — знай это.
Делать
не догадывайся, что твой огонь погас — знай это.
ЛАГЕРЯ. Используйте специально отведенные кемпинги. Держать кемпинги чистые. Горючий мусор сжигают на кострах и весь остальной мусор и отходы всех видов должны быть помещены в мусор банки или ямы, предусмотренные для этой цели. Мертвая или упавшая древесина может быть использована для дрова.
МУСОР.—Не выбрасывайте бумагу, обеденный мусор, кодак. картонные коробки, бумага от жевательной резинки или другой мусор на ободе или на прогулочных дорожках, тропы, дороги или что-то еще.Носите, пока не сможете сгореть в лагере или месте в приемнике.
АВТОМОБИЛЬНЫЕ СРЕДСТВА. Осторожное вождение обязательно.
раз для защиты себя и других посетителей. Твоя машина
должны быть оснащены хорошими тормозами, звуковым сигналом и светом. Переход по кривым
запрещено. Соблюдайте правила дорожного движения. Тракторы с проушинами или транспортные средства
без шин строго запрещены.
PARK RANGERS. Рейнджеры здесь, чтобы помочь и советую вам. Если вы сомневаетесь, спросите рейнджера.Рейнджеры в штаб-квартире парка будем рады помочь вам спланировать свою деятельность в Пещере Ветра и объясните правила.
КАМЕРЫ.— Фото- и кинокамеры могут свободно используется в общих сценических целях.
ПЕЩЕРА ВЕТРОВ ОТКРЫТА ДЛЯ ПУБЛИЧНОСТИ
КАЖДЫЙ ДЕНЬ В ТЕЧЕНИЕ ГОДА.
1940/wica/sec1.htm
Последнее обновление: 20 июня 2010 г.
Апрель Выставка — Галерея Dedee Shattuck
ЗАЯВЛЕНИЕ ХУДОЖНИКА
Sacred Geographies
Я получил приглашение принять участие в выставке Solastalgia как раз в тот момент, когда ущелье реки Колумбия было подожжено.Воздух над Портлендом уже несколько недель был густым от дыма от пожаров, полыхающих от Канады до Калифорнии, окрашивающих луну в зловещий оранжево-красный цвет, покрывающих все пеплом и заставляющих тех из нас, у кого нет кондиционера, держать окна закрытыми, несмотря на 30-градусную жару. понижение температуры ночью, что позволило бы нам охладить наши перегретые дома. Но этот пожар имел гораздо более глубокое воздействие, потому что пожар в Игл-Крик поразил нас в самое сердце. Для большого сообщества Портленд-Ванкувер ущелье реки Колумбия является и домом, и убежищем.Он переносит нас, заземляет нас, возвышает нас, определяет нас. А Игл-Крик был самым сердцем Ущелья. Дом первого кемпинга в Национальной лесной системе с 1916 года, Игл-Крик был типичным местом ущелья, предлагая захватывающие дух виды и культовый водопад Панчбоул. Но теперь безрассудные действия импульсивного подростка угрожали нашему убежищу и нашей коллективной идентичности. Я дважды пролетел над огнем за первые две недели, пока он горел. На этот раз вид, который во всех предыдущих случаях сначала вызывал у меня грусть при уходе, а затем радушно приветствовал меня дома, теперь был окутан дымом.Все, о чем я мог думать, это то, что мы никогда не будем прежними.
понижение температуры ночью, что позволило бы нам охладить наши перегретые дома. Но этот пожар имел гораздо более глубокое воздействие, потому что пожар в Игл-Крик поразил нас в самое сердце. Для большого сообщества Портленд-Ванкувер ущелье реки Колумбия является и домом, и убежищем.Он переносит нас, заземляет нас, возвышает нас, определяет нас. А Игл-Крик был самым сердцем Ущелья. Дом первого кемпинга в Национальной лесной системе с 1916 года, Игл-Крик был типичным местом ущелья, предлагая захватывающие дух виды и культовый водопад Панчбоул. Но теперь безрассудные действия импульсивного подростка угрожали нашему убежищу и нашей коллективной идентичности. Я дважды пролетел над огнем за первые две недели, пока он горел. На этот раз вид, который во всех предыдущих случаях сначала вызывал у меня грусть при уходе, а затем радушно приветствовал меня дома, теперь был окутан дымом.Все, о чем я мог думать, это то, что мы никогда не будем прежними.
Однако после того, как первоначальный шок прошел, массовое излияние коллективного горя, захлестнувшее сообщество, стало казаться почти потакающим своим желаниям. Кто мы такие, чтобы жалеть себя из-за этой потери, и это была действительно огромная потеря, когда так много других людей по всему побережью потеряли свои дома, средства к существованию и даже свои жизни? Да, мы потеряли, возможно, самый драгоценный камень, по крайней мере временно, но уж точно не всю корону драгоценностей.И независимо от того, как он начался, огонь был естественным явлением. Как быстро напомнили нам ученые, на самом деле это было бы хорошо для здоровья леса, важное событие перезагрузки.
Кто мы такие, чтобы жалеть себя из-за этой потери, и это была действительно огромная потеря, когда так много других людей по всему побережью потеряли свои дома, средства к существованию и даже свои жизни? Да, мы потеряли, возможно, самый драгоценный камень, по крайней мере временно, но уж точно не всю корону драгоценностей.И независимо от того, как он начался, огонь был естественным явлением. Как быстро напомнили нам ученые, на самом деле это было бы хорошо для здоровья леса, важное событие перезагрузки.
В то же время уровень язвительности в адрес невольного поджигателя казался столь же неискренним. Самодовольное осуждение очень важного, но необдуманного поступка не учитывало нашу коллективную ответственность и вред, который мы, как некоренное сообщество, наносили этому удивительному месту на протяжении более 150 лет.По крайней мере, с тех пор, как в 1881 году были построены первые железные дороги, мы как общество и наш образ жизни представляли наибольшую угрозу для этого почитаемого места. Наши сложные отношения с ущельем реки Колумбия изменили экосистемы, разрушили традиционные культуры и оставили бесчисленные шрамы на ландшафте из-за лесозаготовок, автомагистралей, плотин, судоходства, ядерных отходов и даже парков, а также недавнего быстрого роста рекреационных походов.
Наши сложные отношения с ущельем реки Колумбия изменили экосистемы, разрушили традиционные культуры и оставили бесчисленные шрамы на ландшафте из-за лесозаготовок, автомагистралей, плотин, судоходства, ядерных отходов и даже парков, а также недавнего быстрого роста рекреационных походов.
Четыре произведения, составляющие серию под названием «Священная география», являются ответом на драматические события осени 2017 года и на противоречивые связи, которые мы поддерживаем даже с нашими самыми священными местами.«Река», «Гора», «Дерево» и «Огонь» отражают красоту мира природы и наши порой необдуманные попытки принадлежать этому миру.
Как пережить сезон змей, даже если вас укусят
25 мая 2018 г. — Добро пожаловать в сезон змей. Когда погода потеплеет, змеи начнут выползать из своих зимних убежищ и, что неизбежно, пересекаются с людьми. Такие встречи могут закончиться поездкой в отделение неотложной помощи.
«В течение весенних и летних месяцев число (посещений скорой помощи, связанных с укусами змей) удваивается, утраивается, а может быть, даже превышает это число», — говорит Дайан Калелло, доктор медицинских наук, доцент кафедры неотложной медицины в Медицинской школе Рутгерса, Нью-Джерси, США. Ньюарк.«Это, безусловно, феномен теплой погоды».
Ньюарк.«Это, безусловно, феномен теплой погоды».
По оценкам CDC, ежегодно в США ядовитые змеи кусают до 8000 человек. Такие укусы редко бывают смертельными. Ежегодно они убивают около пяти человек. Но змеиный яд может нанести серьезный вред.
«Яд вызывает боль и отек и, возможно, образование волдырей в месте укуса, а также повреждает ткани», — говорит Брет Никс, доктор медицинских наук, профессор неотложной медицины в Wake Forest Baptist Health в Уинстон-Салеме, Северная Каролина. «Большие количества могут вызвать внутреннее кровотечение.
В США ядовитые змеи делятся на два типа: гадюки и коралловые змеи.
Ямковидные гадюки имеют много общих физических черт, таких как треугольные головы, вертикальные зрачки и ямка между ноздрями и глазами. Змеи в этой категории включают:
- Гремучие змеи: в США обитает множество видов гремучих змей. Информационно-образовательная система по ядам Джерси.
- Медноголовые змеи: Эти ядовитые змеи живут в основном на востоке США, но их среда обитания простирается на запад до Техаса.
 «Если вас укусит ядовитая змея, вам нужна именно эта, потому что она мало что для вас делает», — говорит Никс.
«Если вас укусит ядовитая змея, вам нужна именно эта, потому что она мало что для вас делает», — говорит Никс. - Вольфрамовые змеи (также известные как водяные мокасины): Виттонмутовые змеи обитают на юго-востоке США. Они любят находиться рядом с водой.
Коралловые змеи, говорит Никс, живут в основном во Флориде и Джорджии. Они черные и красные с белыми или желтыми полосами.
Змеиный яд
На долю гадюк приходится гораздо больше укусов, чем на коралловых змей, но укусы любой из них не всегда ядовиты. Когда они есть, количество яда может варьироваться. Это связано с тем, что змеи контролируют выброс своего яда: чем большую угрозу они воспринимают, тем опаснее их укус. Никс говорит, что каждый четвертый укус гадюки «сухой» или не содержит яда, в то время как 50% укусов коралловых змей доставляют яд.
Вы узнаете, что вас отравила гадюка, если вокруг места укуса появится боль и опухоль.По словам Калелло, змеи чаще всего грызут лодыжку или руку, и симптомы распространяются вверх по ноге или плечу соответственно.
Яд гадюки содержит множество различных соединений, говорит она. Вместе они разрушают ткани и расщепляют белки, которые позволяют вашей крови сворачиваться. Как повреждение тканей, так и кровотечение способствуют отеку.
«Отек и разрушение могут произойти в течение часа, — говорит Калелло, — но повреждение, как правило, не является постоянным, если его лечить.”
При отсутствии лечения повреждение тканей будет ухудшаться. Это может привести к потере конечности или пальцев, говорит Никс, представитель Американского колледжа врачей скорой помощи.
«Если у вас достаточно яда, чтобы вызвать большое количество повреждений тканей, он может стать гангренозным, когда ткань отмирает», — говорит он.
Укусы коралловых змей имеют несколько непосредственных признаков того, что вы были отравлены. До появления серьезных симптомов может пройти от 12 до 18 часов. Яд, содержащий нейротоксины, влияет на способность мозга контролировать мышцы тела.Ваша речь может стать невнятной, и вам может быть трудно глотать. Вы также можете получить слабые мышцы, помутнение зрения и паралич. В конечном итоге яд может затруднить дыхание. Это может привести к летальному исходу, но с 1960-х годов была зарегистрирована только одна смерть от коралловой змеи. Укусы этих змей случаются не часто. Во Флориде, где коралловые змеи наиболее распространены, токсикологические центры ежегодно принимают менее 50 случаев.
Вы также можете получить слабые мышцы, помутнение зрения и паралич. В конечном итоге яд может затруднить дыхание. Это может привести к летальному исходу, но с 1960-х годов была зарегистрирована только одна смерть от коралловой змеи. Укусы этих змей случаются не часто. Во Флориде, где коралловые змеи наиболее распространены, токсикологические центры ежегодно принимают менее 50 случаев.
Передовой опыт
Что делать — и чего не делать — если вас укусила змея:
- DO Как можно скорее обратитесь в отделение неотложной помощи.Даже если укус не ядовит, вы рискуете заболеть столбняком, серьезной бактериальной инфекцией, если ваша прививка не сделана вовремя.
- DO Как можно меньше двигайте конечностью в месте укуса. Это поможет замедлить распространение яда.
- DO Обратите внимание на внешний вид змеи. Ваше описание поможет врачу определить, что вас укусило. (Ваше местоположение также будет подсказкой.
 )
) - DO Запишите время укуса, чтобы врач знал, сколько времени прошло.
- DO Снимите все украшения с места укуса до того, как начнется отек.
- DO При необходимости используйте Тайленол от боли.
- DO Держите под рукой контактную информацию токсикологической службы. Позвоните по номеру 800-222-1222 или отправьте сообщение POISON на номер 797979, чтобы сохранить номер в своем телефоне.
- НЕ ДЕЛАТЬ Пытаться поймать или убить змею. Вы рискуете получить второй укус.
- НЕ ДЕЛАТЬ Попытаться высосать яд. Это не работает, говорит Калелло, и подвергает вас риску попадания яда в рот.
- НЕ ДЕЛАТЬ Принимайте аспирин, ибупрофен или другие болеутоляющие средства, разжижающие кровь.
- НЕ ДЕЛАТЬ Наложить жгут. По словам Калелло, перекрытие притока крови к месту укуса может привести к большему повреждению тканей.

Лечение
Лечение укуса змеи зависит от вида змеи и степени вашей реакции на яд. В легких случаях может потребоваться только очистка места укуса и наблюдение, чтобы убедиться, что вы не причините дальнейшего вреда.Возможно, вам также возьмут кровь, чтобы проверить наличие проблем со свертываемостью крови, вызванных ядом.
В дополнение к первой помощи, наблюдению и анализам крови, лечение более серьезных укусов требует противоядия, которое нейтрализует действие змеиного яда, чтобы ваша кровь могла нормально сворачиваться.
«Существующее противоядие действительно отлично справляется с уменьшением боли и отека в краткосрочной перспективе и улучшением долгосрочных результатов», — говорит Никс.
Противоядие от коралловой змеи также весьма эффективно, но его больше не производят, а запасы истощаются.По данным FDA, текущий запас противоядия коралловой змеи истекает 31 января 2019 г. вспомогательное дыхание. Никс говорит, что отделение неотложной помощи должно координировать свои действия с местным токсикологическим центром, чтобы выяснить, доступно ли противоядие. К счастью, укусы коралловых змей случаются редко.
К счастью, укусы коралловых змей случаются редко.
Противоядие может вызвать немедленную аллергическую реакцию, поэтому вы будете находиться под наблюдением по мере его получения.У некоторых пациентов наблюдается отсроченная реакция, называемая сывороточной болезнью, до 3 недель после введения противоядия. Симптомы включают лихорадку, крапивницу, зуд и боль в суставах.
Ожидайте от 6 до 24 часов наблюдения в больнице, говорит Никс. И будьте готовы к дискомфорту после ухода.
«Симптомы могут длиться в среднем от 1 до 2 недель после лечения», — говорит Калелло.
Профилактика
Конечно, лучше вообще избегать укусов змей, и вы можете сделать это, не провоцируя их.
«Змеи, найденные в США, не агрессивны и кусают только тогда, когда чувствуют угрозу», — говорит Никс. — Они не ищут тебя.
Никс и Калелло рекомендуют следующее в стране змей:
- Обращайте внимание на то, куда вы идете. Змея, скорее всего, не укусит, если вы не наступите на нее.

- Избегайте употребления алкоголя, чтобы сохранять бдительность. (И умно, добавляет Никс: «Выпивка может привести к тому, что вы схватите змею и развернете ее».)
- Берите с собой трость, чтобы постукивать по земле перед собой.Если змея почувствует угрозу, она с большей вероятностью нападет на палку.
- Носите ботинки и длинные брюки, чтобы защитить лодыжки.
- Ночью наденьте налобный фонарь или фонарик, чтобы увидеть змей до того, как вас ждет неожиданная встреча.
%PDF-1.4
%
49 0 объект
>>>/BBox[0 0 531 657]/длина 120>>поток
хА
0DlB=A.jk$oyfpdFAP
А%еҰКНɧɧ==}jYl»7
m*c)l3]O}Il~8
конечный поток
эндообъект
25 0 объект
>>>/BBox[0 0 531 657]/длина 120>>поток
хА
0D1B=А.͋U;ccJ|gU)O$fCpM~œeE)\\~|Igom−~#I
конечный поток
эндообъект
33 0 объект
>>>/BBox[0 0 531 657]/длина 120>>поток
хА
0DlB=A.jk$oyfpdFAP
А%еҰКНɧɧ==}jYl»7
m*c)l3]O}Il~8
конечный поток
эндообъект
51 0 объект
>>>/BBox[0 0 531 657]/длина 120>>поток
хА
0DlB=A.jk$oyfpdFAP
А%еҰКНɧɧ==}jYl»7
m*c)l3]O}Il~8
конечный поток
эндообъект
37 0 объект
>>>/BBox[0 0 531 657]/длина 120>>поток
хА
0DlB=A.